1.炭素循環モデル、炭素循環・気候変化結合モデル3 研究結果の詳細報告へ戻る | HOMEへ戻る |
||||||||||||||
1—3 陸域炭素循環モデルにおける植生帯移動予測モデルの構築担当機関: 地球環境フロンティア研究センター
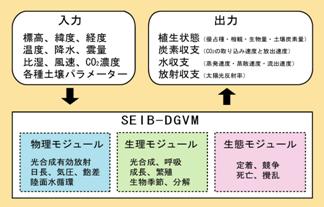
a.要約陸面生態系機能を、数百年から数千年といった時間スケールにおいてシミュレートするためには、気候変動に伴う植生分布までを考慮に入れた動的全球植生モデル(DGVM)が必要とされる。本サブグループの目標は、独自にDGVMを開発し、そのパフォーマンスを十分に検証した後に地球統合モデルに結合させる事である。平成17年度は、前年度までに開発を終えたSEIB-DGVMの更なる改良と、その性能チェックを行った。これまでの試行計算においてSEIB-DGVMは、植物生態系において典型的に観察される幾つかの現象、また現在の気候条件における植生分布や生態系機能分布を再現することに、ある程度まで成功した。 b.研究目的植物生態系の構造(植物種の分布や構成など)と機能(炭素循環や水循環)は気候環境によって強く規定されるが、生態系の構造と機能もまた、蒸発散・炭素循環・陸面粗度の変化・アルベドの変化などを通じて、気候環境にフィードバック的な影響を与える(Foley et al. 2003)。このような気候-植生間の相互作用は、地球環境問題の顕在化に伴い、その定量的解明がますます強く望まれている。気候環境が植生の構造と機能へ与える影響をシミュレートするため、これまで多くのBiogeochemicalモデルが開発され(Peng 2000; Arora 2002)、それらの幾つかはGCM(General Circulation Model)と双方向に結合することで気候-植生間のフィードバックを扱っている(e.g. Woodward et al., 1998; Cox et al., 2000; Joos et al., 2001)。 Biogeochemicalモデルは、静的モデルと動的モデルの2つに大きく区分する事ができる。静的なBiogeochemicalモデル(e.g. Neilson, 1995; Woodward et al., 1995; Haxeltine and Prentice, 1996)は、入力した気候環境の元で光合成・呼吸・成長などの生理過程をシミュレートし、NPPやLAIなどを最大とする植生タイプを優占させる。その際に静的モデルは、気候変化に対して植物生態系の分布が直ぐに平衡に達することを仮定している。しかし、実際には気候が変化しても、その新しい環境に適応した植物生態系が生じるまでには100年~数千年のオーダーの時間遅れがあると予想される(Neilson, 1995; Woodward et al., 1995; Haxeltine and Prentice, 1996)。 そこで、現在地球が経験しているような急速に進行する気候変化の元における、植物生態系の過渡的変化を的確にシミュレートするため、動的なBiogeochemicalモデルが構築されてきた。いずれの動的Biogeochemicalモデルも基本構造はほぼ共通で、既存の静的モデルに植生動態モジュール(定着・競争・死亡・攪乱を扱う)を結合させることで、気候変化に対する植生の過渡的反応のシミュレートを可能にしている(Cramer et al., 2001)。動的モジュールの構造は様々であるが、大きな地理スケールにおける長期演算を可能とするために、いずれも極めて簡便な方式を採用している。例えばLPJモデル(Sitch et al. 2003)では、植物種を10種類のPFTs(Plant Functional Types)で代表させ、グリッド毎に与えられる各PFTの被覆割合を各PFTの単位面積あたりNPPに応じて変化させている。幾つかのモデルでは、PFT間の競争をより機構的に扱ってはいるものの、その殆どは各PFTを平均的な1個体で代表させており(An average individual approach)、PFT内の個体間競争は無視されている。 本研究の目的は、より機構的な植生動態モジュールを有するSEIB-DGVM (Spatialy-Explicit Individual-Base Dynamic-Global-Vegetaion-Model)を新たに構築することである。SEIBは、グリッドボックスごとに幾つかの代表森林(草地)をおき、その中で個体ベースで扱われた木本が定着し、成長し、そして死亡する。このようなモデルの構造は、従来の動的Biogeochemicalモデルに対して次のような利点を持つ。(1) 局所的な光条件によって規定される木本個体間の競争が適切に表現され、したがって気候変動に伴った植生変動の速度を、より的確に予測できることが期待される。(2) ギャップの再生速度が適切に表現され、そのようなギャップ動態に伴う炭素収支の変動(Shugart, 1998)を適切にシミュレートできる。(3) 既存の植物個体群動態の知見やデータとの親和性が高く、パラメーターの推定やモデルの検証が、容易かつ直感的である。このような、局所的競争に基づく植生動態はギャップモデルにより長く解析が行われてきており(Bugmann, 2001)、このような扱いになしには、植生遷移や生産構造を機構ベースで適切に表現できない事が示されている。しかし、ギャップモデルは特定地域の森林動態の再現を目的とする事が多く、これまで全球規模のモデルでは用いられることはなかった。その理由は、より多くの計算力を要求することと、特定の生態系に特異的な相互作用を扱う傾向があるため、より多くのパラメーターを必要とするためである。我々は、最新の計算環境を用い、入手の容易なパラメーターだけで組み上げた植生動態モジュールを開発することにより、これらの制約を克服した。 c.研究計画、方法、スケジュールSEIB-DGVMの基本設計 SEIB-DGVMは、気象・土壌データを入力に用いて、植生の短期的応答(光合成量や呼吸量など)と長期的応答(生物量や生態系の分布など)の両者を出力する(図14)。従来のDGVMsと比較して、SEIB-DGVMを特徴づけているのは、グリッドボックスごとに幾つかの代表森林(草地)をおき、その中で個体ベースで扱われた木本が定着し、成長し、そして死亡する点である(図15)。定着した場所から移動することの出来ない植物にとって、例えば多少の気温上昇よりも、隣の木が枯れて光環境が改善される事の方が、よほど大きな環境変化であり、このような局所的に生じる個体間相互作用を無視しては過渡的な植生変化を的確に予測する事はできない、というのがこの設計を採用した理由である。また、このようなモデルの設計は、既存の植物個体群動態の知見やデータとの親和性が高く、パラメーターの推定やモデルの検証が、容易かつ直感的であるという利点も併せ持つ。このようなアプローチは膨大な計算を伴うため、地球シミュレーターでの運用を前提として初めて可能となった。
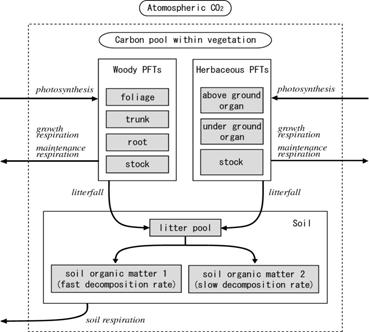
本モデルでは、陸上高等植物種を少数の植物機能型(Plant functional types、以下PFTs)で要約した。この分類法はLPJ-DGVMの分類法に準じ、次の10種類のPFTsを仮定した:熱帯性常緑広葉樹、熱帯性落葉広葉樹、温帯性常緑針葉樹、温帯性常緑広葉樹、温帯性落葉広葉樹、寒帯性常緑針葉樹、寒帯性常緑広葉樹、寒帯性落葉広葉樹、C3草本、C4草本。これらのうち木本PFTは個体ベースで扱われ、各木本個体は樹冠、幹、細根の3部位から構成される。このうち、樹冠と幹は円柱で近似される形態を持つが、細根はバイオマスのみで表現した。他方、草本PFTは葉と根からのみ構成され、それぞれ単位面積あたりのバイオマスによって表現される。 モデル全体の炭素の流れを図16に示す。光合成によって大気中より取り込まれ同化されたCO2は、植物の各器官へ配分される。各器官における維持呼吸や成長呼吸に伴って、ある割合の同化産物は、再びCO2として大気中に放出される。各器官のTurnover、落葉、そして木本の死に伴って生成されるリターは、土壌中のリタープールへと追加される。リターが分解されると、その一部はCO2として大気中に放出され、残りが土壌有機炭素として残留する。この土壌有機炭素は、分解速度の速い画分と遅い画分とに分けられ、これらは分解されるとCO2として大気中に放出される。
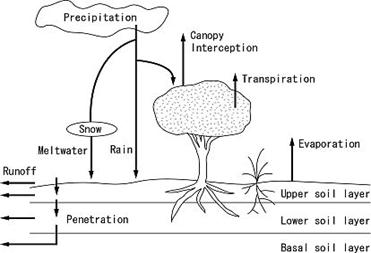
水循環の概略を図17に示す。このモデルでは降水が唯一の水の供給源であり、そして供給された水は「積雪」「土壌上層貯留水」「土壌下層貯留水」のいずれかの形態で仮想林分内に貯留することができる。仮想林分の外に排出される際には、「降雨遮断」「流出」「蒸発」「蒸散」いずれかの経路を通じる。これら一連の水循環過程は、気温・飽差・日射量・土壌粒度などの物理的環境の他、葉面積指数や日光合成量などといった生態的な特徴からも強く制御を受け、そして、土壌貯留水の大小が光合成速度を制御するというフィードバックが働く。
d.平成17年度研究計画平成17年度は昨年度に引き続き、(1)SEIB-DGVMの改良、(2)現在の気象条件を用いた各種シミュレーションによる評価、(3)これらの結果を発表する論文の執筆、を中心として作業を行った。 e.平成17年度研究成果-1.5乗則の検証 小個体が高密度で存在する個体群においては、平均個体重の対数値と個体密度の対数値との間に、通常傾き-3/2の直線関係が成立し、これは-3/2乗則と呼ばれている(Yoda et al. 1963)。この現象は、個体重wは植物の一片lの3乗に比例するのに対し、占有面積a(すなわち光を含む利用可能な資源量)は植物の一片lの2乗に比例する事に起因すると説明されている。すなわち
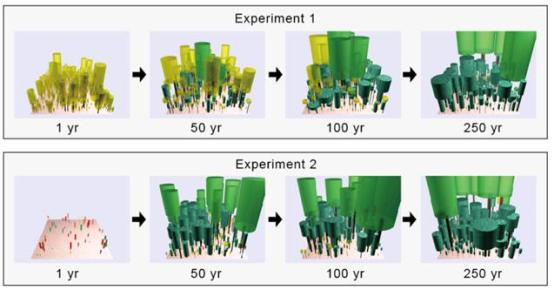
遷移パターンへの初期条件依存性 SEIB-DGVMの最大の特色は、空間構造を明示的に取り扱った仮想林分において、個体ベースで扱われた木本の動態をシミュレートする点にある。これによって、高密度な森林においては既存木本が太陽光を遮り、また定着場所を占め続けることにより、新しい気候環境に適応した木本の侵入が遅れる様子がシミュレートができるはずである。そこで、このような、既存植生の差による環境変動時の植生変化速度への影響が表現されるかどうか、計算実験を行った。シミュレーションは、屋久島サイト(北緯30度20分、年平均気温℃、年降水量3200mm、標高600m、自然植生は温帯性常緑広葉樹林)の気象データを与え、定着可能な木本PFTの間で林床ポイントを等しく分けた。シミュレーションにはパラメーター推定時と同じ気象データを入力に用いたが、スピンアップは次の2通りの条件で行った;処理1. 寒冷条件(元の気象データより、10℃低い気温と地温を用いる)、 処理2. 乾燥条件(元の気象データより、降水量を10分の1とする)。上記の条件で、それぞれ1000年のスピンアップした後、250年間のシミュレーションを行った。 図19に各処理における、シミュレーション開始時点(スピンアップ完了時点)、シミュレーション50年後、シミュレーション100年後、シミュレーション250年後の相観を示した。シミュレーション開始時点においては、処理1は温帯性針葉樹(TeNE)が優占する森林、処理2はC3草本が優占する温帯草原となった(草本は可視化していない)。シミュレーション250年後には、いずれの処理においても温帯性落葉広葉樹(TeBE)とTeBSが優占する森林へと変化するが、処理1ではTeNEの高木が長い期間残存することによって、この植生変化が大幅に遅れた。このような処理間の差は、バイオマス構成の時系列変化を比較すると更に明確であった。例えば、TeBEとTeBSの合計バイオマスが100 Mg C/ haに達するまでの所要期間は、実験1では90年程度、実験2では40年程度であり、そして200 Mg C/ haに達するまでの所要期間は、実験1では190年程度であったが、実験2では110年程度であった。また、実験1ではTeNEのバイオマスが、150年程度も高い状態に留まっていた。これは、気候変化によってpotential vegetationが変化した場合でも、光条件に恵まれた高木は長く留まる事ができる表している。なお、このような既存木本がもたらす植生変化のタイムラグは、新しい環境に適応した木本の種子の制限により増幅されると考えられており(Kohyama & Shigesada 1995)、現実の状況では更に大きいはずである。
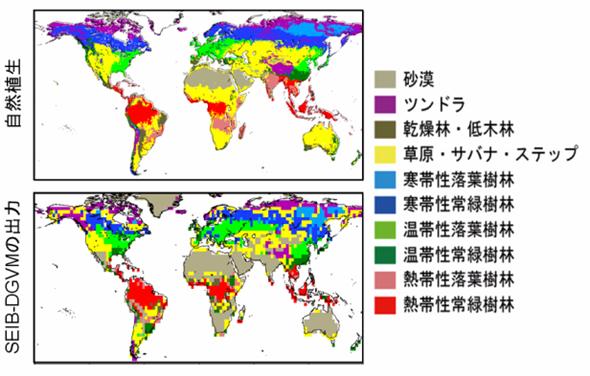
全球実験 T42(128×64 points)の各グリッドを、30m×30mの林分1つで代表させることにより、全球シミュレーションを行った。初期条件は裸地とし、シミュレーション期間は200年間とした。木本PFTの定着条件は次の通りである:最初の50年間は、稚樹が定着できるメッシュを、その場所で定着可能な全ての木本PFTへ等しく分配させ、50年目以降は現存バイオマスに比例して分配させる。図20は、植生の分布と自然植生とを、シミュレーション200年後との間の比較である。自然植生の分布図は、Haxeltine and Prentice (1996)を簡素化した。また、シミュレーション出力の植生タイプの分類基準も、Haxeltine and Prentice (1996)の基準を簡素化したものを用いた(Table 10)。なお、このnatural vegetation mapの作成には、植生データのみが用いられ、気象データや他のモデルの出力は用いられていない。 全体的に、SEIB-DGVMは全球の植生分布を、ある程度までは再現することに成功したといえる。しかし、幾つかの地域の植生タイプ、特に熱帯域の乾燥の程度に応じた植生タイプの移り変わりなどは、適切に再現されなかった。例えば、インド全域やアフリカのサハラ以南には、自然植生では熱帯季節林・サバナ・Xeric woodlandといった乾燥環境に適応した熱帯性植生が生じているが、シミュレーションでは砂漠が出力さてしまっている。
f.考察一連のシミュレーション結果は、SEIB-DGVMが植生動態・炭素循環・陸面水循環の概略を、少なくとも幾つかの選択地点においては再現できることを明らかにした。また、既存植生の違いが、植生の移行速度へ影響する様子を、特別なパラメタリーゼーションなしで表現することが出来た。これは、本モデルの特徴的な設計(個体ベース+明示的な空間構造)によって初めて可能となったことであり、従来のDGVMsでは為しえなかったことである。今後は、温暖化やCO2濃度の増加がおきた際の生態系機能や植生分布の変化パターンを調べ、これらを既存のモデルの結果と比較することにより、本モデルの特徴的な設計が、モデル出力にどのような与えるのか更に検討したい。また我々の研究プロジェクトでは、現在開発が進められている地球統合モデルKISSME(Kawamiya et al. 2005)へSEIB-DGVMを結合させ、陸面-大気間の相互作用が今後の地球環境変動にどのような影響を与えうるのかシミュレーション実験を行う予定である。 SEIB-DGVMは、既存のどの全球植生モデルよりも実際の植生動態メカニズムを明示的に反映させているものの、幾つかの問題点が残存することを指摘しておく。第1には全ての陸上維管束植物を僅か10種類のPFTでまとめている点である。全球シミュレーションにおいて、自然植生を正しく再現できなかった地域が生じた理由の一つは、この多様性の単純化に端を発している可能性が高いと考える。例えば、本モデルではxeric woodland/scrubが生じなかったが、そのようなバイオームで優占種となる灌木種をモデルに導入する事で解決できると期待される。 問題点の第2は、木本PFTの定着率を一定としている点である。木本の定着は栄養繁殖能、種子休眠性、耐陰性等に大きく影響を受け(reviewed by Greene et al. 1999)、そのような定着の可否は、植生動態や景観を強く規定している。しかし、定着率は様々な要因によって決まる複雑かつ多様な過程であり、これを全球モデルにおいて機構ベースで取り込むことは現実的ではないだろう。一つの可能性としては、各木本PFTの定着率を遷各移段階や各気候区分別に経験則として与えてやることである。Starfield & Chapin (1996)は、そのような経験則をベースとしたモデルALFRESCOにおいて、アラスカの過渡的な景観変化パターンを、気候条件・山火事からの経過年数・森林伐採の頻度などの関数として再現することに成功している。 問題点の第3は、広大な地域を30m×30mといった小さな平面空間で代表させてしまい、景観の多様性を考慮に入れていない点である。従来の高度に単純化されたモデルでは、そのような景観の多様性を含めてパラメタライズしていることが暗に想定されており、この点には大きな注意が払われてこなかった。しかし、実際の機構をより強くモデルに反映させるに従って、この問題の相対的な重要性も増すであろう。ある限定された地域のシミュレーションであれば、単純にグリッドメッシュを細かくし、各グリッドメッシュに個別の環境条件を与えてやるというのが本質的な解決法となる。しかし、全球計算のためには、計算量を節約するため、何らかのsophisticateされた手法を模索しなければならない。これも今後の課題である。 シミュレーションモデルの構造を複雑することは、モデル出力の不確実性を増大させる。したがって、上記で指摘した様々なプロセスを次々と結合していくことには、十分な注意が必要である。しかし、気候変動時の植生応答を予測するという、実験のできない現象を高い精度で予測するためには、その現象において本質的な過程は、少なくとも簡素化した形では導入しなければならない。そして、陸面生態系プロセスは複雑かつ多様であるため、それは多くの生態学者の協力を仰がなければ為しえない事業である。SEIB-DGVMは、このような陸域生態学者の知見を集積させる過程において、中心的な役割を担うことが出来る。なぜならば、SEIB-DGVMは、木本の定着・競争・死亡といった局所的空間構造に制約されらながら個体ベースで生じる現象を明示的にシミュレートする、唯一の全球モデルだからである。問題となるのはSEIBが大きな計算力を要求する点である。Hurtt et al.(1998)やMoorcroft et al.(2001)は、確率的なデモグラフィー過程を近似する方法を提案しており、これは、そのような過程を限られた計算力で扱うことを可能にする。しかし、SEIB-DGVMは一林分における計算であれば小型のワークステーションで十分に運用可能であり、また計算環境は今後ますます充実していく方向にある。さらに、明示的な計算は明解性や直感性が高いことから、幅広い分野の研究者から知見を集めるのに適している。 なおSEIB-DGVMは、さらなる改良を加えることでモデル信頼性の向上に努め、農業生態系や土地利用変化モジュールを導入した後に、共生第2プロジェクトにおいて開発が進行している地球統合モデルへと結合される事が予定されている。 g.引用文献Arora, V., 2002. Modeling vegetation as a dynamic component in soil-vegetation-atmosphere transfer schemes and hydrological models. Rev. Geophys. 40, 1006, doi:10.1029/2001RG000103. Bugmann, H., 2001. A review of forest gap models. Clim. Change 51, 259-305. Cox, P.M., Betts, R.A., Jones, C.D., Spall, S.A., Totterdell, I.J., 2000. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature 408, 184-187. Cramer, W., Bondeau, A., Woodward, F.I., Prentice, I.C., Betts, R.A., Brovkin, V., Cox, P.M., Fisher, V., Foley, J.A., Friend, A.D., Kucharik, C., Lomas, M.R., Ramankutty, N., Sitch, S., Smith, B., White, A., Young-Molling, C., 2001. Global response of terrestrial ecosystem structure and function to CO2 and climate change: results from six dynamic global vegetation models. Global Change Biol 7, 357-373. Foley, J.A., Costa, M.H., Delire, C., Ramankutty, N., Snyder, P., 2003. Green surprise? How terrestrial ecosystems could affect earth's climate. Frontier Ecol. Environ. 1, 38-44. Greene, D.F., Zasada, J.C., Sirois, L., Kneeshaw, D., Morin, H., Charron, I., Simard, M.-J., 1999. A review of the regeneration dynamics of North American boreal forest tree species. Can. J. For. Res. 29, 824-839. Haxeltine, A., Prentice, I.C., 1996. BIOME3: An equilibrium terrestrial biosphere model based on ecophysiological constrains, resource availability, and competition among plant functional types. Global Biogeochem. Cycles 10, 693-709. Hurtt, G.C., Moorcroft, P.R. Pacala, S.W., and Levin, S.A., 1998. Terrestrial models and global change: Challenges for the future. Global Change Biology 4, 581-590. Joos, F., Prentice, I.C., Sitch, S., Meyer, R., Hooss, G., Plattner, G., Gerber, S., Hasselmann, K., 2001. Global warming feedbacks on terrestrial carbon uptake under the Intergovernmental Panel on Climate Change (IPCC) emission scenarios. Global Biogeochem. Cycles 15, 891-907. Kawamiya, M., Yoshikawa, C., Sato, H., Sudo, K., Watanabe, S., Matsuno, T., 2005. Development of an Integrated Earth System Model on the Earth Simulator. Journal of Earth Simurator 4, 18-30. Kohyama, T., Shigesada, N., 1995. A size-distribution-based model of forest dynamics along a latitudinal environmental gradient. Vegetatio 121, 117-126. Kohyama, T., 2005. Scaling up from shifting-gap mosaic to geographic distribution in the modeling of forest dynamics. Ecol. Res. 20, 305-312. Moorcroft, P.R., Hurtt, G.C., and Pacala, S.W., 2001. A method for scaling vegetation dynamics: The ecosystem demography model (ED). Ecological Monographs 71, 557-586. Neilson, R.P., 1995. A model for predicting continental-scale vegetation distribution and water balance. Ecol. Appl. 5, 362-385. Peng, C.H., 2000. From static biogeographical model to dynamic global vegetation model: a global perspective on modelling vegetation dynamics. Ecol. Modell. 135, 33-54. Shugart, H.H., 1998. The mosaic theory of natural landscapes. In: Terrestrial ecosystems in changing environments. Cambridge University Press, pp. 178-204. Sitch, S., Smith, B., Prentice, I.C., Arneth, A., Bondeau, A., Cramer, W., Kaplan, J.O., Levis, S., Lucht, W., Sykes, M.T., Thonicke, K., Venevski, S., 2003. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Global Change Biol. 9, 161-185. Starfield, A.M., Chapin III, F.S., 1996. Model of transient changes in arctic and boreal vegetation in response to climate and land use change. Ecol. Appl. 6, 842-864. Woodward, F.I., Lomas, M.R., Betts, R.A., 1998. Vegetation-climate feedbacks in a greenhouse world. Phil. Trans. R. Soc. Land. B 353, 29-39. Woodward, F.I., Smith, T.M., Emanuel, W.R., 1995. A global land primary productivity and phytogeography model. Global Biogeochem. Cycles 9, 471-490. Yoda, K., Kira, T., Ogawa, H., Hozumi, K., 1963. Self-thinning in overcrowded pure stands under cultivated and natural conditions (Intraspecific competition among higher plants XI). Journal of Biology, Osaka City University 14, 107-129. h.成果の発表<口頭発表> Sato, H., A. Itoh, and T. Kohyama, SEIB-DGVM: A Spatial-Explicit Individual-Base Dynamic-Global-Vegetation-Model, ESA-INTECOL 2005 joint meeting, Montreal, 2005. Sato, H., A. Itoh, and T. Kohyama, SEIB-DGVM: A Spatial-Explicit Individual-Base Dynamic-Global-Vegetation-Model, 11th US-Japan Workshop on Global Change, Yokohama, 2005.(ポスター) Sato, H., A. Itoh, and T. Kohyama, SEIB-DGVM: A Spatial-Explicit Individual-Base Dynamic-Global-Vegetation-Model, 11th US-Japan Workshop on Global Change, Yokohama, 2005.(ポスター) Sato, H., A. Itoh, and T. Kohyama, SEIB-DGVM: A Spatial-Explicit Individual-Base Dynamic-Global-Vegetation-Model, 1st iLEAPS Science Conference, Boulder USA, 2006.(ポスター) Sato, H., A. Itoh, and T. Kohyama, SEIB-DGVM: A Spatial-Explicit Individual-Base Dynamic-Global-Vegetation-Model, The 8th International Workshop on Next Generation Climate Models for Advanced High Performance Computing Facilities, Albuquerque USA, 2006. 佐藤永, 伊藤明彦, 甲山隆司, SEIB-DGVM: A Spatial-Explicit Individual-Base Dynamic-Global-Vegetation-Model, 南西諸島における森林生態系研究に関する現状と展望, 琉球大学農学部亜熱帯フィールド科学教育研究センター, 2005. 佐藤永, 伊藤明彦, 甲山隆司, SEIB-DGVM: A Spatial-Explicit Individual-Base Dynamic-Global-Vegetation-Model, 日本気象学会2005年秋季大会, 神戸大学, 2005. 佐藤永, 伊藤明彦, 甲山隆司, SEIB-DGVM: A Spatial-Explicit Individual-Base Dynamic-Global-Vegetation-Model, 横浜国立大学オープンセミナー, 横浜国立大学, 2005. 佐藤永, 伊藤明彦, 甲山隆司, SEIB-DGVM: A Spatial-Explicit Individual-Base Dynamic-Global-Vegetation-Model, 第53回日本生態学会大会, 新潟コンベンションセンター, 2006. <論文出版> Sato, H., A. Itoh, and T. Kohyama, SEIB-DGVM: A New Dynamic Global Vegetation Model using a Spatially Explicit Individual-Based Approach, Submitted to Ecological Modelling. Kawamiya, M., C. Yoshikawa, H. Sato, K. Sudo, S. Watanabe, and T. Matsuno, Development of an Integrated Earth System Model on the Earth Simulator, Journal of Earth Simurator, 4, 18-30, 2005. 次のページ(2.1 温暖化・大気組成変化相互作用(大気化学)) |

 と
と より、
より、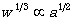 となり、
となり、 となる。そして、一個体の専有面積aは個体密度dと反比例の関係(
となる。そして、一個体の専有面積aは個体密度dと反比例の関係( )にあるため、
)にあるため、 という関係が導き出される。そこで本モデルにおいて、この関係が成立するか確かめるための計算実験を行った。寒帯・温帯・熱帯の3地域に、それぞれ稚樹を初期密度1(tree/m2)で配置し、100年間のシミュレーションを行った。この100年間の間には新しい稚樹の定着はさせなかった。図18は、このシミュレーションにおける木本密度と平均個体重との関係の変化を示した両対数グラフである。稚樹がある程度成長した以降は、いずれの地域においても概ね傾き-3/2の関係で両者が変化し、-3/2乗則が成立した。これより、樹木の専有面積とバイオマスとの相対成長関係が、本モデルにおいて適切に再現されていると言うことが出来る。
という関係が導き出される。そこで本モデルにおいて、この関係が成立するか確かめるための計算実験を行った。寒帯・温帯・熱帯の3地域に、それぞれ稚樹を初期密度1(tree/m2)で配置し、100年間のシミュレーションを行った。この100年間の間には新しい稚樹の定着はさせなかった。図18は、このシミュレーションにおける木本密度と平均個体重との関係の変化を示した両対数グラフである。稚樹がある程度成長した以降は、いずれの地域においても概ね傾き-3/2の関係で両者が変化し、-3/2乗則が成立した。これより、樹木の専有面積とバイオマスとの相対成長関係が、本モデルにおいて適切に再現されていると言うことが出来る。