
成果
[2004年-2010年]
未培養系統群好熱性アーキアゲノムを解明
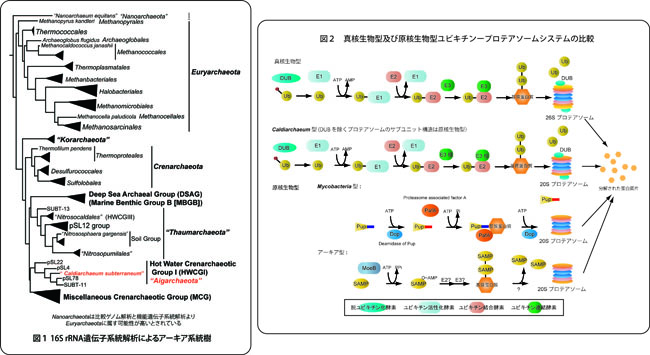
シュガープログラムはゲノムチーム、京都大学 跡見晴幸教授、慶応大学 金井昭夫教授、東京大学 服部正平教授のグループと共同で、地下鉱山の熱水に繁茂する微生物マットのメタゲノム解析を行い、誰もが培養したことのない系統に属する好熱性アーキア(恐らく70度程度で増殖する。Caldiarchaeum subterraneumと名付けた。'熱いアーキア''地殻内生物'の意)の全ゲノム解析に成功しました(図1参照)。
勿論、シュガープログラムのワシは1031(世界のKT?)も、このアーキアの培養にはアッサリと跳ね返されています。共生菌を除けば、培養できない微生物の全ゲノム解析に成功したのは日本国内では初めてです。世界でも、共生菌や集積培養を経ていない試料からの例に限れば、両手の指に収まるくらいの報告しかありません。色んな"巡り会い"なしには、自然環境中の試料から、特定微生物の全ゲノム解析なんて出来ませんでした。良い試料・共同研究者・技術・研究資金その他諸々、色んなものが上手くはまりました。このプロジェクト自体、そもそもはCaldiarchaeum と共存する別のアーキア(2008年にアメリカで集積培養されたアンモニア酸化好熱性アーキア;ちなみに1031はこのアーキアの培養にも至らず。)のゲノムを解析しようと始めた仕事です。それがあれよあれよという間に別のアーキア(Caldiarchaeum )の全ゲノム解析にまで成功してしまったのです。2004年に始めた仕事ですから(数度の中断もあり時間がかかりました)、本来の目的の通りに行っていれば2008年の好熱性アンモニア酸化アーキア集積培養成功!で半泣きになっていたはず。しかも、Caldiarchaeum を含む試料を採取できたのは、同じフィールドで何度もサンプリングしていますが、後にも先にも、このメタゲノム解析試料のみ。ですので、どんな形をしている細胞かも分かっていません。紆余曲折を経ましたが、ホント、'何か持ってます'。
さて、アーキアのゲノムを構成する遺伝子を既知のアーキアゲノムと比較しますと、このアーキアは、既存のCrenarchaeota門、Euryarchaeaota門は勿論、最近提唱されているThaumarchaeota門、Korarchaeota門とも異なる特徴を示しました。つまり、このアーキアを含む系統群が新たな門を構成する可能性が示されたのです。そこでAigarchaeota門(夜明け・オーロラアーキアの意)をシレッと提唱しています。ここまではアーキアオタクの話題。バイオロジーのグローバルな観点から見て最も重要なことは、真核生物と原核生物を分ける特徴とされてきた、真核生物型のユビキチン修飾に関する遺伝子が見つかったことです。この発見はちょっとしたセレンディピティですね。そもそも、このメタゲノム解析は、別のアーキアの生理生態を知りたくて始めたプロジェクトですし、Caldiarchaeum の生理生態は'フーン、ただの好熱性アーキアだったんだ・・・'と大して感激もありませんでしたから、これがなければ、アーキアの世界だけの話題でした。ユビキチン修飾系の代表的なものは、ユビキチンという小さな蛋白質で不要な蛋白質を標識し、その標識を目印にプロテアソームが不要な蛋白質を分解する仕組みです(図2参照)。最近、アーキア型あるいはマイコバクテリア(結核菌の類です)型と呼ばれる類似のシステムは発見されていましたが、やっぱり原核生物から見つかるものは真核生物型の仕組みとは違うね(所詮、下等よね)といった感じで片付けられていました。
今回の発見は、真核生物とアーキアの共通祖先が既に真核生物型のユビキチンシステムを持っていたこと、大部分の?アーキアが何故か真核生物型のユビキチンシステムを放棄したことを意味します。実際の機能的な証明は、今しばらくお待ち下さい。ゲノムから想像した絵が証明できれば勝ちです。また、このメタゲノム解析には、論文でも触れていない分子生物学上の結構重要な発見がいくつかあり、共同研究者が解析中です。まだ秘密ですが、お楽しみに。
- このトピックに関するプレスリリースは、こちらから