Profile

Marine Ecosystem Research Group
Senior Researcher
Address
Japan Agency for Marine-Earth Science and Technology (JAMSTEC)
Earth Surface System Research Center (ESS)
2-15 Natsushima, Yokosuka, Kanagawa, 237-0061, Japan
Education
- Ph.D., Water Resources Program, 1998 Civil Engineering, Princeton University
- B.S., Chemical Engineering, 1992 California Institute of Technology (Caltech)
Vertical migration by bulk phytoplankton sustains biodiversity and nutrient input to the surface ocean
Kai Wirtz1 and S. Lan Smith
Scientific Reports, vol. 10, January 24, 2020
Phytoplankton subsumes the great variety of unicellular photoautotrophs that perform roughly half of Earth’s primary production. They achieve this despite their challenging oceanic habitat, with opposing vertical gradients of nutrients (which often limit their growth near the surface) and light (which becomes limiting with increasing depth). Most phyto- plankton species are commonly assumed to be incapable of moving actively between the zones of light and nutrient availability, which are separated vertically by from 30–120m. Here we propose that a considerable fraction of phyto- plankton vertically traverse these gradients over time scales from hours to weeks, employing variations of a common migration strategy to acquire multiple resources. We present a mechanistic Lagrangian model resolving phytoplank- ton growth linked to optimal migration behaviour and demonstrate unprecedented agreement of its calculated vertical CHL-a distributions with 773 profiles observed at five prominent marine time-series stations. Our simulations reveal that vertically cycling phytoplankton can pump up enough nutrient to sustain as much as half of oceanic Net Primary Production (NPP). Active locomotion is therefore a plausible mechanism enabling relatively high NPP in the olig- otrophic surface ocean. Our simulations also predict similar fitness for a variety of very different migration strategies, which helps to explain the puzzling diversity of phytoplankton observed in the ocean.

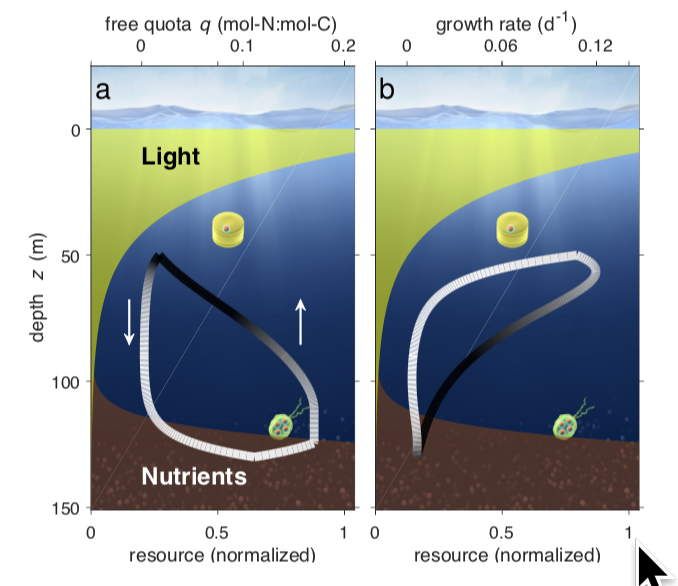
Micro-scale patchiness enhances trophic transfer efficiency and potential plankton biodiversity
Anupam Priyadarshi, S. Lan Smith, Sandip Mandal, Mamoru Tanaka & Hidekatsu Yamazaki
Scientific Reports volume 9, Article number: 17243 (2019)
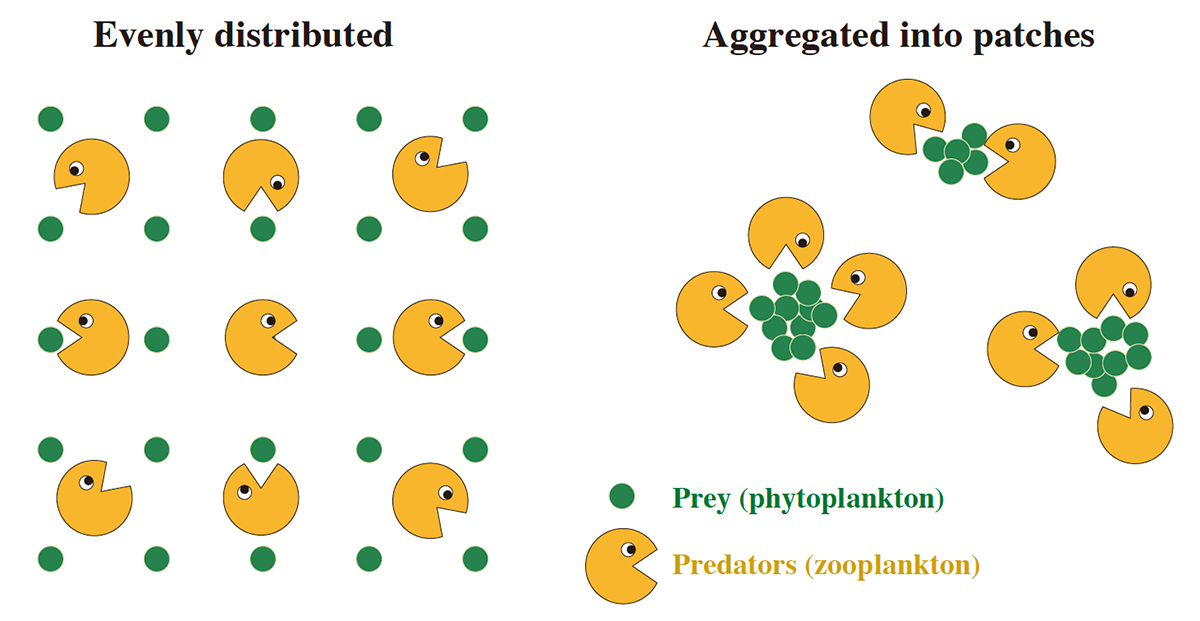
Rather than spatial means of biomass, observed overlap in the intermittent spatial distributions of aquatic predators and prey is known to be more important for determining the flow of nutrients and energy up the food chain. A few previous studies have separately suggested that such intermittency enhances phytoplankton growth and trophic transfer to sustain zooplankton and ultimately fisheries. Recent observations have revealed that phytoplankton distributions display consistently high degrees of mm scale patchiness, increasing along a gradient from estuarine to open ocean waters. Using a generalized framework of plankton ecosystem models with different trophic configurations, each accounting for this intermittency, we show that it consistently enhances trophic transfer efficiency (TE), i.e. the transfer of energy up the food chain, and expands the model stability domain. Our results provide a new explanation for observation-based estimates of unexpectedly high TE in the vast oligotrophic ocean and suggest that by enhancing the viable trait space, micro-scale variability may potentially sustain plankton biodiversity.
Effect of phytoplankton size diversity on primary productivity in the North Pacific: trait distributions under environmental variability. by Bngzhang Chen, S. Lan Smith, and Kai W. Wirtz. Ecology Letters vol. 22, 2019, doi: 10.1111/ele.13167."
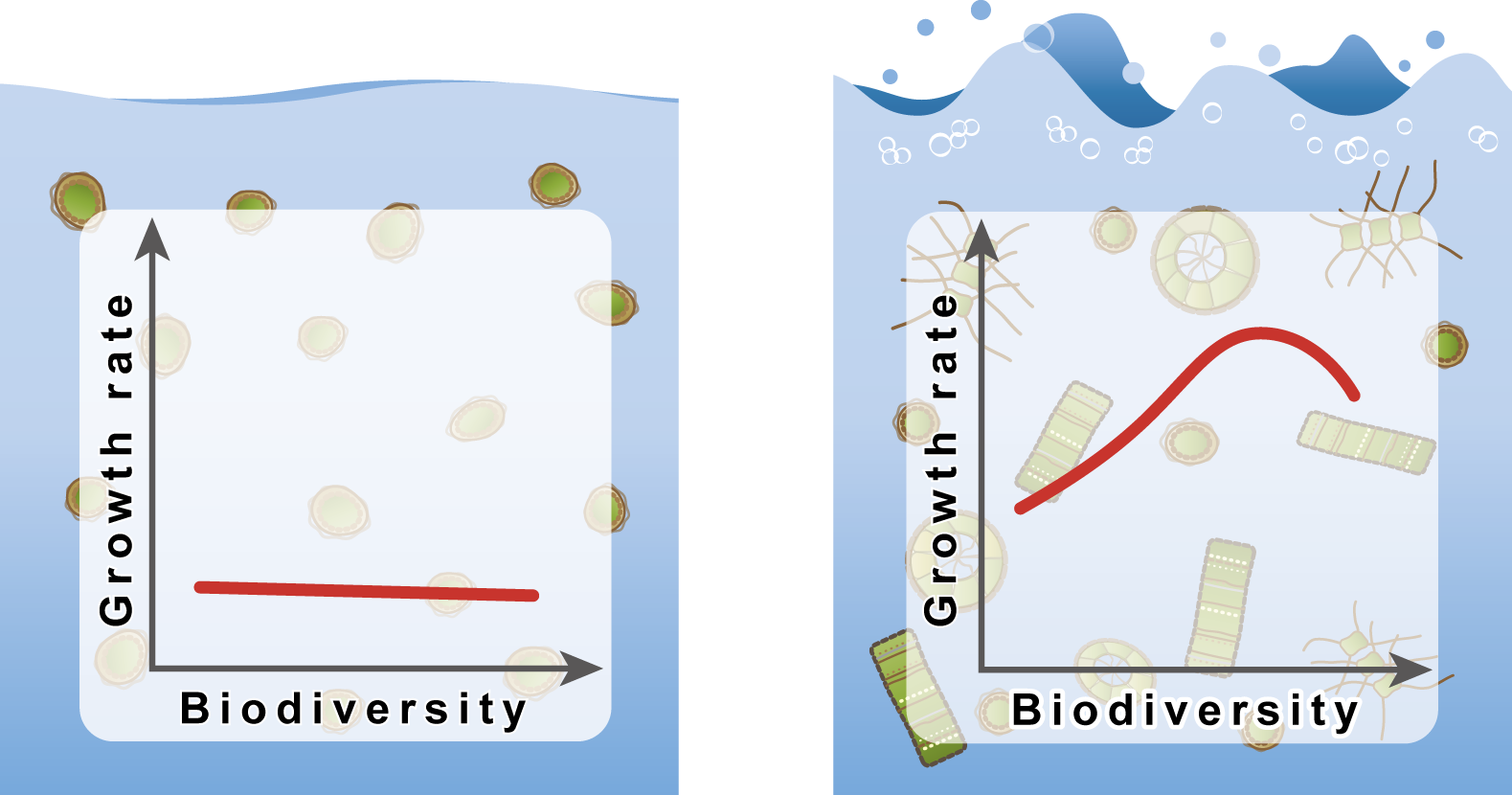
Phytoplankton size-diversity mediates an emergent trade-off in ecosystem functioning for rare versus frequent disturbances
S. Lan Smith, Sergio M. Vallina & Agostino Merico
Scientific Reports, vol. 6, 17 October, 2016
In environments with less frequent disturbance, phytoplankton communities with less diversity tend to be slightly more productive (left). On the contrary, when disturbances are more frequent, more diverse communities tend to be more productive.
Related Links
Research theme :
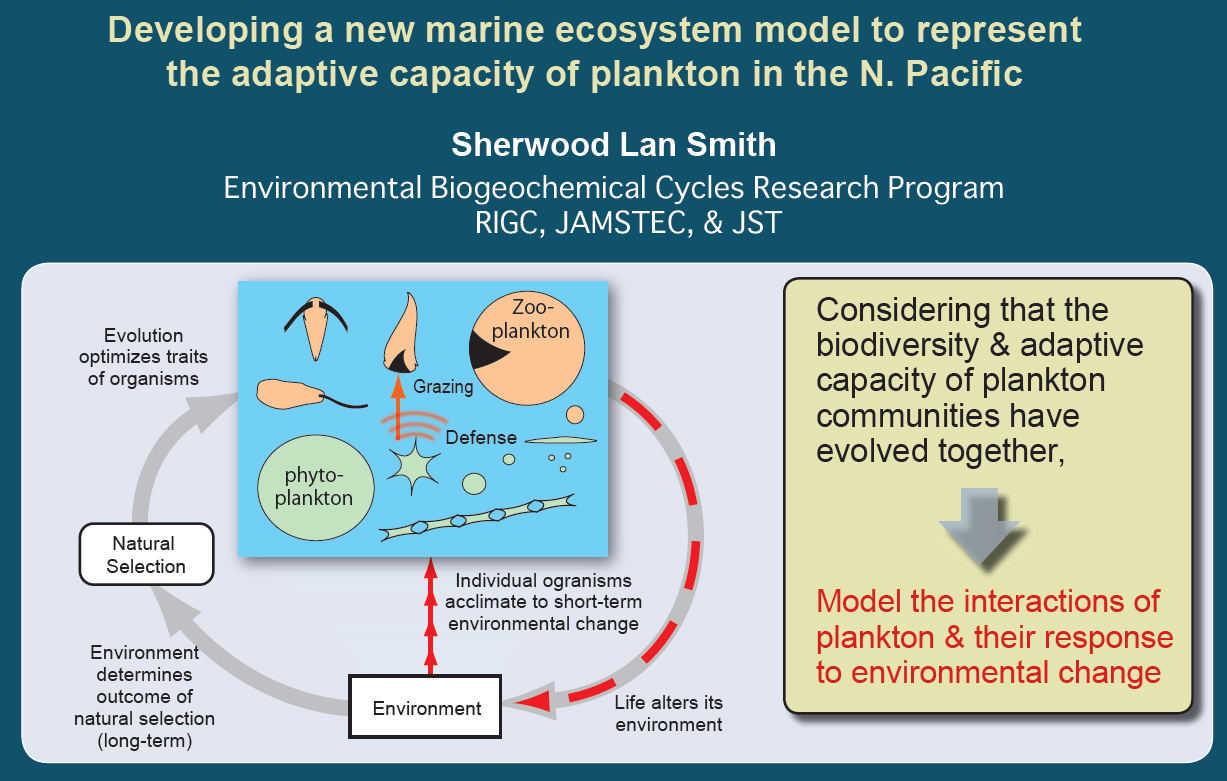
Interpreting observations and developing better models of plankton based on the concept of Optimality (as a result of natural selection)
Developing a new marine ecosystem model to represent the adaptive capacity of plankton in the N. Pacific
CREST (Core Research for Evolutional Science and Technology) Project funded by JST (Japan Science and Technology Agency), FY2012 to FY2017
We intend to develop the new ecosystem model based on the 'adaptive dynamics' approach, using trade-offs to constrain the adaptive response.
Related Links
Introduction to Optimality-based modeling
Natural selection tends to produce organisms optimally adapted to their environments.
Optimality can be considered a guiding concept.
Particularly for plankton:
- long evolutionary history
- short generation times
- easy to use in experiments
Many studies have made progress by assuming:
Adaptation (species) & Acclimation (organisms) both tend to maximize growth rate.
- Simpler models,
- Models of greater generality.
- New, different interpretations of observations.
Solve the Problem faced by organisms:
How to make the best use of limited resources, to maximize fitness*, subject to trade-offs in a variable environment?
*For asexually reproducing plankton, we equate fitness with growth rate.
Successful applications at various levels of organization.
Ideally, net growth rate should be optimized, but this is only possible for simple systems.
However, for many natural systems, this is not practical because it requires modeling the entire ecosystem, including predators and all other losses, which are not well known.
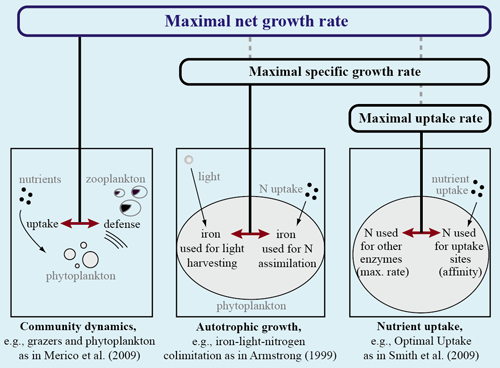
Nevertheless, the optimality concept has been applied successfully to model lower levels of organization, e.g., the cellular level or specific processes such as uptake.
This makes sense for sub-processes which are vital for growth.
Red arrows show the Trade-offs, linked to the quantity optimized (above).
Selected Examples
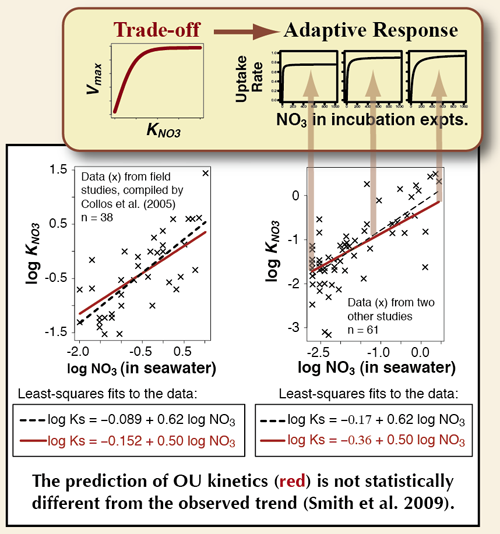
Optimal Uptake (OU) kinetics
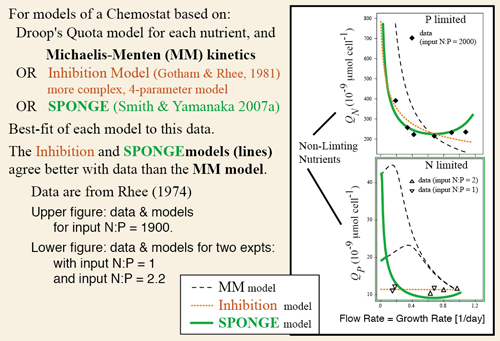
1. agrees better with observations of nutrient uptake by phytoplankton and bacteria, compared to the widely applied Michaelis-Menten (MM) equation (Smith & Yamanaka 2007a; Smith et al. 2009, 2011),
2. reproduces observed changes in Si and N uptake during the SERIES iron-enrichment expt. (Smith et al. 2010), and
3. provides a more consistent interpretation of the combined effects of temperature and concentration on nitrate uptake as observed on oceanic cruises, and implications for the C cycle (Smith 2010, 2011).
Photoacclimation models
- 4. agree better with observations over wider ranges of environmental conditions, compared to previous models, despite having fewer parameters (Pahlow 2005; Smith & Yamanaka 2007b; Pahlow & Oschlies 2009; Wirtz & Pahlow 2010),
- 5. provide a new and more general interpretation of co-limitation of phytoplankton by N and P (Pahlow and Oschlies 2009), and
- 6. reproduce the shape of the often observed “Droop curve” for growth rate from basic principles, i.e., without specifying a particular functional form (Wirtz and Pahlow 2010).
Selected Results
Selected results from 1.
Selected results from 3.
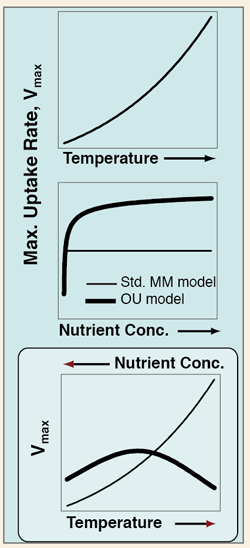
With OU kinetics, Vmax (as measured by short-term expts.) depends on ambient [NO3] as well as on T. This results in greater inferred T sensitivity, compared to the standard assumption of T dependence only (as applied with Michaelis-Menten kinetics).
Compared to MM kinetics, with OU kinetics greater T sensitivity is required to reproduce the same field data, because as T increases [NO3] tends to decrease (right).
For a data set for nitrate uptake from the N. Atlantic, Smith (2011) found
Q10 ~ 3 based on OU kinetics
as great as for heterotrophic bacteria
vs.
Q10 ~ 2 based on MM kinetics
lower than for heterotrophic bacteria
![Because T and [NO3] are negatively related in the near-surface ocean (as shown above for the data set of Harrison et al. (1996) from the N. Atlantic, the effects of T and concentration tend to counteract one another for Vmax.](images/fig4.png)
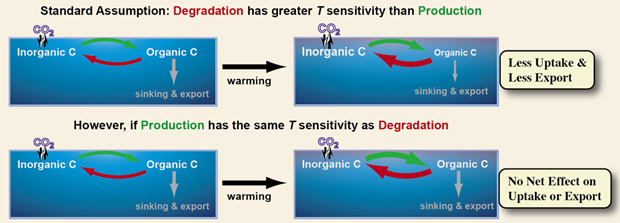
Implications for the carbon cycle
Conclusions
The concept of optimality is quite valuable for modeling plankton.
Still, important challenges remain, including:
-
Practical:
Obtaining observations of sufficient quality and quantity to rigorously test competing theoretical models. -
Theoretical:
How to develop useful dynamic models based on optimality that are also consistent with the theory of the Evolutionarily Stable Strategy (ESS, by John Maynard Smith), which is formulated in terms of steady state?
References


Flexible phytoplankton functional type (FlexPFT) model: size-scaling of traits and optimal growth
S. Lan Smith, Markus Pahlow, Agostino Merico, Esteban Acevedo-Trejos, Yoshikazu Sasai, Chisato Yoshikawa, Kosei Sasaoka, Tetsuichi Fujiki, Kazuhiko Matsumoto and Makio C. Honda
J. Plankton Res. (2015) doi: 10.1093/plankt/fbv038 First published online: June 3, 2015
 Leaving misleading legacies behind in plankton ecosystem modelling
Leaving misleading legacies behind in plankton ecosystem modelling
S. Lan Smith; Agostino Merico; Kai W. Wirtz; Markus Pahlow
Journal of Plankton Research 2014; doi: 10.1093/plankt/fbu011
- Armstrong, RA (1999) An optimization-based model of iron–light–ammonium colimitation of nitrate uptake. Limnology & Oceanography 44, p. 1436–1446
- Collos Y, A Vaquer and P Souchu (2005) Acclimation of nitrate uptake by phytoplankton to high substrate levels. Journal of Phycology 41, p. 466-478
- Harrison, GW, LR Harris, and DB Irwin (1996), The kinetics of nitrogen utilization in the oceanic mixed layer: Nitrate and ammonium interactions at nanomolar concentrations. Limnology & Oceanography 41, p. 16–32
- Merico A, J Bruggeman and K Wirtz (2009) A trait-based approach for downscaling complexity in plankton ecosystem models. Ecological Modelling 220, p. 3001-3010
- Pahlow M (2005) Linking chlorophyll-nutrient dynamics to the Redfield N:C ratio with a model of optimal phytoplankton growth. Marine Ecology Progress Series 287, p. 33-43
- Pahlow M and A Oschlies (2009) Chain model of phytoplankton P, N and light colimitation. Marine Ecology Prog. Ser. 376, p. 69-83
- Rhee, G-Y (1974) Phosphate uptake under nitrate limitation by Scenedesmus sp. and its ecological implications. Journal of Phycology 10, p. 470–475
- Smith SL (2010) Untangling the uncertainties about the combined effects of temperature and concentration on nitrate uptake in the ocean. Geophysical Research Letters 37 L11603, doi:10.1029/2010GL043617
- Smith SL (2011) Consistently modeling the combined effects of temperature and concentration on nitrate uptake in the ocean. J. Geophysical Research--Biogeosciences 116, G04020, doi:10.1029/2011JG001681
- Smith SL, M Pahlow, A Merico and KW Wirtz (2011) Optimality-based modelling of planktonic organisms. Limnology & Oceanography 56 Review paper, p. 2080-2094
- Smith SL and Y Yamanaka (2007a) Optimization-based model of multinutrient uptake kinetics. Limnology & Oceanography 52, p. 1545-1558
- Smith SL and Y Yamanaka (2007b) Quantitative comparison of photoacclimation models for marine phytoplankton. Ecological Modelling 201, p. 547-552
- Smith SL, Y Yamanaka, M Pahlow and A Oschlies (2009) Optimal Uptake Kinetics: Physiological acclimation explains the observed pattern of nitrate uptake by phytoplankton in the ocean. Marine Ecology Progress Series 384 Feature Article, p. 1-12
- Smith SL, N Yoshie and Y Yamanaka (2010) Physiological acclimation by phytoplankton explains observed changes in Si and N uptake rates during the SERIES iron-enrichment expt. Deep Sea Res. I 57, p. 394-408
- Wirtz, KW and M Pahlow (2010) Dynamic chlorophyll and nitrogen:carbon regulation in algae optimizes instantaneous growth rate. Marine Ecology Progress Series 402, p. 81-96
Presentations
Invited Public Lecture at JpGU2019 annual meeting, Makuhari Messe, Chiba (May 25, 2019).
This session was co-organized by two JAMSTEC Scientists (Drs. Yoshikawa & Ohgaito) for the benefit of Japanese scientists who must present their research results in English, with particular emphasis on students and early career scientists as the intended audience.

Publications
- Anupam Priyadarshi, S. Lan Smith, Sandip Mandal, Mamoru Tanaka, and Hidekatsu Yamazaki (2019). Micro-scale patchiness enhances trophic transfer efficiency and potential plankton biodiversity. Scientific Reports, vol. 9, doi: 10.1038/s41598-019-53592-6
- Sandip Mandal, S. Lan Smith, Anupam Priyadarshi, and Hidekatsu Yamazaki (2019) Micro-scale variability impacts the outcome of competition between different modelled size classes of phytoplankton. Frontiers in Marine Science, section Marine Ecosystem Ecology.doi: doi.org/10.3389/fmars.2019.00259
- S. Lan Smith, Sandip Mandal, Anupam Priyadarshi, Bingzhang Chen, and Hidekatsu Yamazaki (2019) Modeling the Combined Effects of Physiological Flexibility and Micro-Scale Variability for Plankton Ecosystem Dynamics. Chapter in Cochran, J. Kirk; Bokuniewicz, J. Henry; Yager, L. Patricia (eds.) Encyclopedia of Ocean Sciences, 3rd Edition, vol.[5], pp. 527-535. Oxford: Elsevier. doi: 10.1016/B978-0-12-409548-9.10955-8
- Bingzhang Chen, S. Lan Smith, and Kai W. Wirtz (2019) Effect of phytoplankton size diversity on primary productivity in the North Pacific: trait distributions under environmental variability. Ecology Letters vol. 22, doi: 10.1111/ele.13167
- Models in Microbial Ecology.
SM Vallina, R Martinez-Garcia, SL Smith, JA Bonachela
Encyclopedia of Microbiology, Reference Module in Life Sciences (Elsevier), 2019 - Optimality-based approach for computationally efficient modeling of phytoplankton growth, chlorophyll-to-carbon, and nitrogen-to-carbon ratios.
B Chen, SL Smith
Ecological Modelling 385, 197-212, 2018 - CITRATE 1.0: Phytoplankton continuous trait-distribution model with one-dimensional physical transport applied to the North Pacific.
B Chen, SL Smith
Geoscientific Model Development 11, 467-495, 2018 - Comparison of carbon cycle between the western Pacific subarctic and subtropical time-series stations: highlights of the K2S1 project.
MC Honda, M Wakita, K Matsumoto, T Fujiki, E Siswanto, K Sasaoka, ...
Journal of Oceanography 73 (5), 647-667, 2017 - Micro-scale variability enhances trophic transfer and potentially sustains biodiversity in plankton ecosystems.
A Priyadarshi, S Mandal, SL Smith, H Yamazaki
Journal of Theoretical Biology 412, 86-93, 2017 - Modeling phytoplankton size structure and diversity using the ‘adaptive dynamics’ approach.
B Chen, SL Smith Kaiyo Monthly 49, 446-463, 2017 - PhytoSFDM version 1.0. 0: phytoplankton size and functional diversity model.
E Acevedo-Trejos, G Brandt, SL Smith, A Merico
Geoscientific Model Development 9 (11), 4071-4085, 2016 - Phytoplankton size-diversity mediates an emergent trade-off in ecosystem functioning for rare versus frequent disturbances.
SL Smith, SM Vallina, A Merico
Scientific Reports (Nature Publishing Group) 6, 34170, 2016 - Introduction to the Themed Section: Advances in plankton modeling and biodiversity evaluation.
H Yamazaki, SL Smith
Journal of Plankton Research 38 (4), 944-945, 2016 - Contribution to the Themed Section: Advances in Plankton Modelling and Biodiversity Evaluation A 1D physical-biological model of the impact of highly intermittent phytoplankton …
S Mandal, H Homma, A Priyadarshi, H Burchard, SL Smith, KW Wirtz, ...
JOURNAL OF PLANKTON RESEARCH 38 (4), 964-976, 2016 - Contribution to the Themed Section: Advances in Plankton Modelling and Biodiversity Evaluation Flexible phytoplankton functional type (FlexPFT) model: size-scaling of traits …
SL Smith, M Pahlow, A Merico, E Acevedo-Trejos, Y Sasai, C Yoshikawa, ...
JOURNAL OF PLANKTON RESEARCH 38 (4), 977-992, 2016 - Coupled 1-D physical–biological model study of phytoplankton production at two contrasting time-series stations in the western North Pacific
Y Sasai, C Yoshikawa, SL Smith, T Hashioka, K Matsumoto, M Wakita, ...
Journal of Oceanography 72 (3), 509-526, 2016 - A 1D physical–biological model of the impact of highly intermittent phytoplankton distributions
S Mandal, H Homma, A Priyadarshi, H Burchard, SL Smith, KW Wirtz, ...
Journal of Plankton Research 38 (4), 964-976, 2017 - Sustaining diversity in trait-based models of phytoplankton communities.
A Merico, G Brandt, SL Smith, M Oliver
Frontiers in Ecology and Evolution 2, 59, 2014 - Sizing-up nutrient uptake kinetics: combining a physiological trade-off with size-scaling of phytoplankton traits.
SL Smith, A Merico, S Hohn, G Brandt
Marine Ecology Progress Series 511, 33-39, 2014 - Leaving misleading legacies behind in plankton ecosystem modelling.
SL Smith, A Merico, KW Wirtz, M Pahlow
Journal of Plankton Research 36 (3), 613-620, 2014 - Relating Phytoplankton Pnysiology to North Pacific Biogeochemistry.
SL Smith, MN Aita, M Shigemitsu, Y Yamanaka
In Western Pacific Air-Sea Interaction Study, eds. Uematsu et al. (TERRAPUB), p. 223-235, 2014 - Biogeographical patterns of phytoplankton community size structure in the oceans.
E Acevedo‐Trejos, G Brandt, A Merico, SL Smith
Global Ecology and Biogeography 22 (9), 1060-1070, 2013 - Temperature‐induced marine export production during glacial period.
MO Chikamoto, A Abe‐Ouchi, A Oka, SL Smith
Geophysical Research Letters 39 (21), 2012 - Development of a one‐dimensional ecosystem model including the iron cycle applied to the Oyashio region, western subarctic Pacific.
M Shigemitsu, T Okunishi, J Nishioka, H Sumata, T Hashioka, MN Aita, ...
Journal of Geophysical Research: Oceans 117 (C6), 2012 - Consistently modeling the combined effects of temperature and concentration on nitrate uptake in the ocean.
SL Smith
Journal of Geophysical Research: Biogeosciences 116 (G4), 2011 - Optimality‐based modeling of planktonic organisms.
SL Smith, M Pahlow, A Merico, KW Wirtz
Limnology and Oceanography 56 (6), 2080-2094, 2011 - Linear relationship between carbon and nitrogen isotope ratios along simple food chains in marine environments.
MN Aita, K Tadokoro, NO Ogawa, F Hyodo, R Ishii, SL Smith, T Saino, ...
Journal of plankton research 33 (11), 1629-1642, 2010 - Untangling the uncertainties about combined effects of temperature and concentration on nutrient uptake rates in the ocean.
SL Smith
Geophysical research letters 37 (11), 2010 - Physiological acclimation by phytoplankton explains observed changes in Si and N uptake rates during the SERIES iron-enrichment experiment.
SL Smith, N Yoshie, Y Yamanaka
Deep Sea Research Part I: Oceanographic Research Papers 57 (3), 394-408, 2010 - End‐to‐end models for the analysis of marine ecosystems: challenges, issues, and next steps.
KA Rose, JI Allen, Y Artioli, M Barange, J Blackford, F Carlotti, R Cropp, ...
Marine and Coastal Fisheries 2 (1), 115-130, 2010 - Optimal uptake kinetics: physiological acclimation explains the pattern of nitrate uptake by phytoplankton in the ocean.
SL Smith, Y Yamanaka, M Pahlow, A Oschlies
Marine Ecology Progress Series, Feature Article 384, 1-12, 2009 - Examining the effects of nutrients on the composition and size of phytoplankton using unaltered deep-sea waters.
MP Niraula, BE Casareto, SL Smith, T Hanai, Y Suzuki
Journal of Experimental Marine Biology and Ecology 348 (1-2), 23-32, 2007 - Optimization‐based model of multinutrient uptake kinetics.
S Lan Smith, Y Yamanaka
Limnology and Oceanography 52 (4), 1545-1558, 2007 - A bioenergetics-based population dynamics model of Pacific herring (Clupea harengus pallasi) coupled to a lower trophic level nutrient–phytoplankton–zooplankton model …
BA Megrey, KA Rose, RA Klumb, DE Hay, FE Werner, DL Eslinger, ...
Ecological Modelling 202 (1-2), 144-164, 2007 - NEMURO—a lower trophic level model for the North Pacific marine ecosystem.
MJ Kishi, M Kashiwai, DM Ware, BA Megrey, DL Eslinger, FE Werner, ... SL Smith, et al.
Ecological Modelling 202 (1-2), 12-25, 2007 - Quantitative comparison of photoacclimation models for marine phytoplankton.
SL Smith, Y Yamanaka
ecological modelling 201 (3-4), 547-552, 2007 - Examining the regeneration of nitrogen by assimilating data from incubations into a multi-element ecosystem model.
SL Smith, BE Casareto, MP Niraula, Y Suzuki, JC Hargreaves, JD Annan, ...
Journal of Marine Systems 64 (1-4), 135-152, 2007 - Attempting consistent simulations of Stn. ALOHA with a multi-element ecosystem model.
SL Smith, Y Yamanaka, MJ Kishi
Journal of oceanography 61 (1), 1-23, 2005 - Development of carbon biomass using incubations of unaltered deep-sea water.
YS Mohan P. Niraula, Beatriz E. Casareto, Takayuki Hanai, S. Lan Smith
Eco-Engineering 17 (2), 121-131, 2005 - The dynamics of arsenic in saturated porous media: fate and transport modeling for deep aquatic sediments, wetland sediments, and groundwater environments.
SL Jaffe, P.R., Wang, S., Kallin, P.L. and Smith
Special Publication 7, 379-397, 2002 - Modeling the transport and reaction of trace metals in water‐saturated soils and sediments
SL Smith, PR Jaffé
Water Resources Research 34 (11), 3135-3147, 1998