プレスリリース
2018年 2月 2日
国立研究開発法人海洋研究開発機構
国立大学法人北海道大学
国立大学法人京都大学
独立行政法人製品評価技術基盤機構
生命誕生に迫る始原的代謝系の発見
~多元的オミクス研究による新奇TCA回路の証明~
1.概要
国立研究開発法人海洋研究開発機構(理事長 平 朝彦、以下「JAMSTEC」という)海洋生命理工学研究開発センターの布浦拓郎主任研究員らは、北海道大学(総長 名和 豊春)、京都大学(総長 山極 壽一)、製品評価技術基盤機構(理事長 辰巳 敬、以下「NITE」という)と共同で、南部沖縄トラフの熱水活動域から採取した試料より単離した細菌が、アミノ酸など生命に必須の化合物の生合成に不可欠なTCA回路(tricarboxylic acid 回路:クエン酸回路)の中でも、最も始原的な形態の回路を有することを発見しました。
多くの生物にとりTCA回路は生存に必須の代謝機構であり、その起源は生命の共通祖先の誕生、更には化学進化の時代にまで遡る「最も始原的な基幹代謝のひとつ」と考えられています(図1)。TCA回路にはいくつかの形態が存在し、生命誕生前後の始原的なTCA回路の姿については様々な議論があります。
今回、布浦主任研究員らの研究グループは、始原的バクテリア系統に属する好熱性水素酸化硫黄還元細菌Thermosulfidibacter takaii(図2、以下「Thermosulfidibacter」という)が最も始原的な形態のTCA回路を持つことを示しました。本研究における多元的オミクス解析(※1)の結果、Thermosulfidibacterが独立栄養(※2)または混合栄養(※3)条件でも、全く同じ酵素群を用い、利用できる炭素源に応じて回路の反応方向を柔軟に変化させる、可逆的なTCA回路を保持していることが明らかになりました(図3)。これまで全く同じ酵素群を用いたTCA回路で独立栄養と従属栄養の両方の機能を使い分ける生物は見つかっていませんでした。Thermosulfidibacterで観察された新奇TCA回路はダイナミックに変動する環境条件に適応して反応の向きを切り替えるという、最も祖先型のTCA回路が備えていたであろう特性を示していると考えられます。
初期生命の形態は、独立栄養であったのか、あるいは従属栄養であったのか、永い議論が続いています(図4)。今回示した、この始原的な新奇TCA回路の特性は、「初期生命が原始地球の生命誕生の場において、利用可能な物質の存在量に応じて、柔軟に代謝を変化させる混合栄養生命として誕生した」可能性を強く示しています。
本研究の一部は、国立研究開発法人科学技術振興機構(JST)の戦略的創造研究推進事業(CREST)の助成を受けて実施されました。
本成果は、米国科学誌「SCIENCE」に2月2日付け(日本時間)で掲載される予定です。
タイトル:A primordial and reversible TCA cycle in a facultatively chemolithoautotrophic thermophile
著者名:布浦拓郎1 力石嘉人1,2,3 井崎力久4 諏訪貴士 4 佐藤喬章4 原田健史5 森浩二5 加藤裕美子5 宮崎征行1,6 島村繁6 柳川勝紀6† 首藤彩1 大河内直彦2 藤田信之5‡ 高木善弘1,6 跡見晴幸4,7 高井研6
所属:1.国立研究開発法人海洋研究開発機構 海洋生命理工学研究開発センター、2. 国立研究開発法人海洋研究開発機構 生物地球化学研究分野、3.北海道大学 低温科学研究所、4.京都大学大学院工学研究科、5.独立行政法人製品評価技術基盤機構、6. 国立研究開発法人海洋研究開発機構深海・地殻内生物圏研究分野、7. 国立研究開発法人科学技術振興機構 CREST
現所属
†北九州市立大学 ‡東京農業大学
2.背景
地球上における生命の誕生時の形態が、自ら炭酸固定を行う独立栄養生物であったのか、あるいは非生物学的に生じた有機物を利用する従属栄養生物であったのか、永い議論が続いています。しかし、いずれの生命形態であったとしても、アミノ酸等の生命維持に不可欠な化合物の生合成の起点となるピルビン酸、アセチルCoA、オキサロ酢酸、オキソグルタル酸、スクシニルCoAの生合成を担うTCA回路(図1)は、生命の誕生時あるいはそれ以前の化学進化の時代においても、不可欠であったと考えられています。その一方、TCA回路は非常に柔軟に形を変える仕組みであることが知られています。実際、回路の一部を欠く生物や変形型を有す生物が存在し、殆どの反応において起源の異なる複数の酵素が知られており、また、回路の反応方向にも多様性が存在します(図5)。
その多様なTCA回路の中でも、還元的TCA回路は、最も古い炭素固定経路である可能性が指摘されています。しかし、還元的TCA回路に必須の酵素(※4)が進化的に新しい起源を有し、生命誕生時には存在しなかった可能性が高いことも議論されてきました。
3.成果
本研究の開始において、JAMSTEC とNITEは共同研究により、好熱性水素酸化硫黄還元細菌Thermosulfidibacter の全ゲノム解析を実施しました。その結果、ゲノムからは完全な形の既知炭素固定経路は確認できず、唯一、クエン酸合成酵素 (通常はオキサロ酢酸とアセチルCoAからクエン酸とCoAを合成する)が逆反応(吸エルゴン反応である)で機能する、新奇な還元的TCA回路が存在する可能性が示されました。そこで、Thermosulfidibacter細胞におけるTCA回路に関わる酵素活性を測定すると共にトランスクリプトーム及びプロテオーム解析を実施しました。更に、JAMSTECにおいて新しく開発された微量メタボローム解析技術を用い、北海道大学とも共同で、各培養条件におけるTCA回路の方向を検討しました。その結果、独立栄養条件下においては還元方向(反時計回り:図1)に回路が機能して炭酸固定を行い、また、混合栄養条件において酢酸やコハク酸を添加した場合、それぞれアセチルCoA、コハク酸を分岐点とする回路が機能して、回路の一部でそれぞれ脱炭酸と炭酸固定を行うことが明らかとなりました。そして、有機物の供給形態によっては、炭酸固定はせず、脱炭酸のみを進めることも示されました(図3)。
更に、JAMSTECと京都大学は共同で、大腸菌で発現したThermosulfidibacterのクエン酸合成酵素について、酵素化学的な解析を実施しました。その結果、Thermosulfidibacterのクエン酸合成酵素は既知のATPクエン酸リアーゼと同程度にオキサロ酢酸とアセチルCoAの合成に適応した性質を有し、還元的TCA回路にも十分に機能しうることが明らかにされました。また、本研究において、改めてクエン酸合成酵素とATPクエン酸リアーゼ、シトリルCoAリアーゼの系統関係を解析し、ATPクエン酸リアーゼやシトリルCoAリアーゼはクエン酸合成酵素の一群より進化した可能性が極めて高いことも確認されました。
これら一連の結果は、Thermosulfidibacterは、異なる増殖条件においてクエン酸合成酵素を含む全く同じ酵素群が機能して、供給される中間代謝物の濃度に対応して柔軟にTCA回路の反応の向きを切り替えていることを示しています。そして、クエン酸合成酵素の逆反応を利用した還元的TCA回路の存在を初めて明らかにしました。
TCA回路は、図1に示す左側の経路は、反時計回りの還元反応において発エルゴン反応(自発反応)であり、右側の反応は時計回りの酸化反応において発エルゴン反応です。よく知られる好気性微生物やミトコンドリアが有する酸化的TCA回路も含め、回路として機能する為には、生物は何らかの仕組みで吸エルゴン反応を進める必要があります。本研究の対象であるThermosulfidibacterは、有機物を利用して増殖する際にも電子供与体である水素と電子受容体である硫黄を必要とします。Thermosulfidibacterでは、この水素を酸化し硫黄を還元するエネルギー代謝で得たエネルギーを上手く利用することで、TCA回路の反応の向きを増殖条件に適した形に制御していると考えられます。
本研究が示すTCA回路の柔軟な性質は、化学エネルギーが十分に供給される熱水環境で誕生した生物が、Thermosulfidibacterと同様、その場に存在する炭酸と有機物種とその量に応じて、独立栄養と混合栄養の生活様式を柔軟に変更していたことを示唆しています(図4)。即ち、生命は従属栄養や独立栄養に特化した生物ではなく、所与の環境条件に柔軟に適応する存在として誕生したと考えられます。
4.今後の展望
始原的なアーキア、バクテリア系統群においても、一部の欠失を含めTCA回路には多様性が存在することが、ゲノム解析より示されています。しかし、これらは既知の生化学的知見とゲノム情報の対照から得られたものであり、実際の反応方向や、代替経路、未知酵素が関連する可能性等は全く検証されていません。生命誕生時のTCA回路の形態や役割に関する理解を深めるには、これらの多様なTCA回路の姿を明らかにすることが不可欠です。
また、今回の解析に用いた新規な微量メタボローム解析技術は、従来の同様の技術で要求される細胞量の百分の一以下で実施可能です。この新規解析技術により、十分な細胞量を得ることができないため、基礎/応用研究の両面におけるバイオリソースとしての利用が極めて困難であった環境微生物における代謝研究が可能となりました。未知生合成経路の探索等、新しい環境微生物利用の道が開けたと言えるでしょう。今後は、従来利用されてこなかった環境微生物からの未知生合成経路の探索や、TCA回路における多様性の理解は、生命の本質に迫るだけでなく、システム生物学における代謝設計にも新たな可能性を提供し、新しいバイオテクノロジー展開にも貢献することが期待されます。
JAMSTECと京都大学は、包括連携協定を有しており、この枠組みを軸に、今後も環境微生物が有す未知微生物代謝系・生合成系の探索等を推進します。JAMSTECとNITEは、深海バイオリソースを産官学におけるイノベーションに繋げるため、今後も連携を進めます。 JAMSTECと北海道大学は、今後も連携して、新規メタボローム解析技術の汎用化と高度化を進めます。
[用語解説]
※1 多元的オミクス研究:ゲノミクス(遺伝子:DNA)、トランスクリプトミクス(遺伝子:RNA)プロテオミクス(蛋白質)、メタボロミクス(代謝物)等を包括して解析・解明する研究のこと。
※2 独立栄養:本発表においては化学合成独立栄養を意味する。Thermosulfidibacterの場合は、水素の還元的化合物を電子供与体として、エネルギーを獲得し、そのエネルギーを利用して無機炭素からの有機物合成を行う。
※3 混合栄養:Thermosulfidibacterにおいては、水素を電子供与体として利用することが増殖に不可欠である。水素を電子供与体として、エネルギーを獲得する一方、取り込んだ有機物を主要な炭素源とする。また、増殖条件により、有機物と無機炭素を共に炭素源として併用する。
※4 還元的TCA回路においては、クエン酸とCoAからオキサロ酢酸とアセチルCoAを合成する反応は吸エルゴン反応(非自発的反応)である。既存の知見では、ATPクエン酸リアーゼ(クエン酸とCoAをATPのエネルギーを用いてオキサロ酢酸とアセチルCoAに変換する)あるいはその代替の仕組み(シトリルCoA合成酵素はクエン酸とCoAからシトリルCoAを合成し、シトリルCoAリアーゼはシトリルCoAをATPのエネルギーを用いてオキサロ酢酸とアセチルCoAに変換する)が還元的TCA回路には不可欠とされていた。
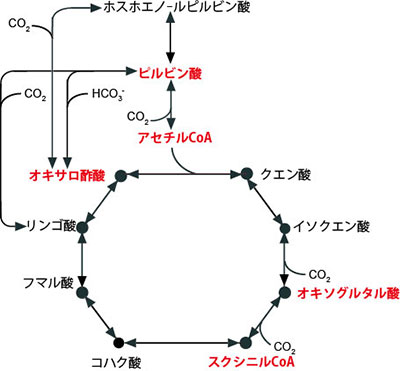
図1. TCA回路とピルビン酸、アセチルCoA、オキサロ酢酸、オキソグルタル酸、スクシニルCoAの関係。
図2. Thermosulfidibacter takaiiの細胞
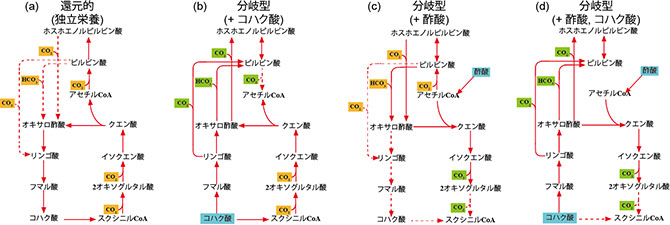
図3.微量メタボローム解析により示されたThermosulfidibacterの可逆的TCA回路。独立栄養条件(a)では還元的TCA回路が、コハク酸(b)、酢酸(c)、両方(d)添加時にはそれぞれ異なる分岐型TCA回路が生じる。コハク酸(b)と酢酸(c)を添加した際では炭酸同化と脱炭酸を行う位置が異なる。また、コハク酸と酢酸の両方を添加した際(d)には脱炭酸のみが生じる。
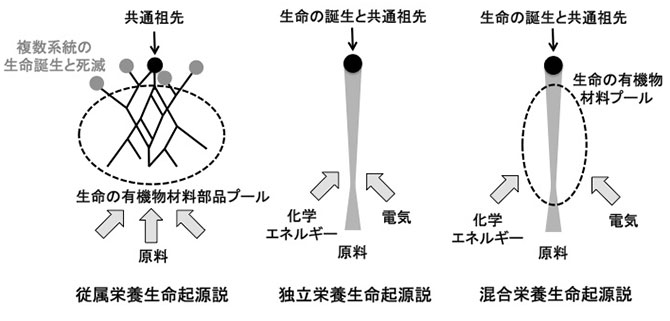
図4.生命誕生のイメージ。有機物プールから誕生したとする従属栄養生命起源説、無機触媒・化学エネルギー等を基盤として生命が誕生したとする独立栄養生命起源説の間で議論が続いている。従属栄養生命起源説では生命活動の永続性に、独立栄養生命起源説では生命活動に必要な濃度に有機物が濃集する仕組みにおいて脆弱性が指摘されている。混合栄養生命起源説であれば、既存説の脆弱性を解決することが可能である。
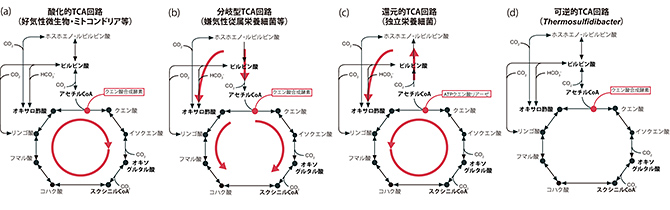
図5.これまでに知られていたTCA回路の主な類型(a-c)と今回Thermosulfidibacterより見出された可逆的TCA回路(d)。
- (本研究について)
- 国立研究開発法人海洋研究開発機構
- 海洋生命理工学研究開発センター 研究開発センター長代理 布浦 拓郎
- (報道担当)
- 広報部 報道課長 野口 剛
- 国立大学法人北海道大学
- 総務企画部 広報課 北野 誉直
- 国立大学法人京都大学
- 総務部 広報課 国際広報室 國包ターヒューン レイモンド
- 独立行政法人製品評価技術基盤機構
- バイオテクノロジーセンター 計画課戦略企画室 市川 夏子