1.炭素循環モデル、炭素循環・気候変化結合モデル3 研究結果の詳細報告へ戻る | HOMEへ戻る |
|||||||||||||||||||||||||||
1—2.海洋生物地球化学モデル担当機関名:地球フロンティア研究システム
a.要約大気海洋結合モデルへの炭素循環過程組み込みの前段階として、海洋単体モデルへ組み込んだ炭素循環モデルを用い、予備的な温暖化実験を行った。すなわち、大気海洋結合モデルを用い過去に行われた二酸化炭素漸増実験の結果得られた風応力、海表面温度を海洋モデルの駆動力として用い、温暖化によって海洋環境が変化した場合とそうでない場合とについて大気中二酸化炭素濃度が漸増したときの海洋二酸化炭素吸収量が両ケースの間でどのように異なるのか調べた。その結果、温暖化による海洋循環変化が二酸化炭素吸収量に与える影響は小さいという結果が得られた。本プロジェクトで採用しているモデルはIPCC第3次報告書で採用されているものよりも海洋表層生態系の記述が詳細になっているが、ここでの温暖化実験では過去の実験結果を追認するに終わった。このこと自体は華やかな結果ではないが、陸域炭素循環と結合させる前段階として、我々のモデルがもっともらしい振る舞いを見せることは確認できたと言える。このことを受け、大気海洋結合モデルへの炭素循環モデル移植にも着手し、現在その原型が完成した段階にある。今後、モデルパラメータのチューニングやコードの整備、充分なスピンアップを行い、炭素循環−気候結合モデル相互比較プロジェクト(C4MIP)への参加、ひいてはIPCC第4次報告書への貢献に備える。さらに、植物・動物プランクトンの種構成を陽に表現したモデルを用いた実験も行い、カイアシ類の季節鉛直移動や10年スケールの気候変動が海洋表層生態系に与える影響などについて調べた。 b.研究目的海洋中の全炭酸鉛直分布は表層付近で濃度が低くなる特徴的な分布をしている。二酸化炭素の大気海洋交換にとって大きな意味を持つこうした分布は生物ポンプ・アルカリポンプ・物理ポンプといった過程によって決定されており、中でも表層生態系における有機物の形成とそれに続く沈降に起因する生物ポンプが最も重要な寄与をなしている。その生物ポンプの効率は、海洋混合層の深さやエクマン湧昇、大気による鉄分の輸送など様々な物理過程から影響を受けている。人間活動により排出された二酸化炭素がどの程度大気中に残存するかを把握し、将来の大気中二酸化炭素濃度の予測を確からしいものにするためには、海洋中の炭素循環過程をきちんとモデル化することが不可欠である。 ハドレ−センター(英)やIPSL(仏)が行った陸域−大気−海洋結合炭素循環モデルの結果によれば、気候変動が海洋の二酸化炭素吸収に与える影響は小さいとされる(Cox et al.,2000; Friedlingstein et al.2001)。しかしながら、海洋炭素循環モデル相互比較プロジェクト(Ocean Carbon-Cycle Model Intercomparison Project,OCMIP)に提出された結果を見ると、気候変動を考慮に入れずに行ったベースラインの海洋二酸化炭素吸収量将来予測において、モデル間のばらつきが大きくなっており、2100年時点での予測値は最小値と最大値の間で2倍の開きがある(Fasham,2003)。大気中二酸化炭素濃度の予測のためには、引き続き海洋炭素循環モデルを改善し、こうした不確定性を減らしていくことが必要である。本研究テーマでは、4変数の単純な海洋生態系モデルを炭素循環モデルとともに海洋大循環モデルへ組み込んで海洋炭素循環と気候変化との相互作用を調べ、さらに発展して陸域−大気−海洋結合炭素循環モデルの構築とそれによる全球規模炭素循環の研究を行うことを目的にしている。
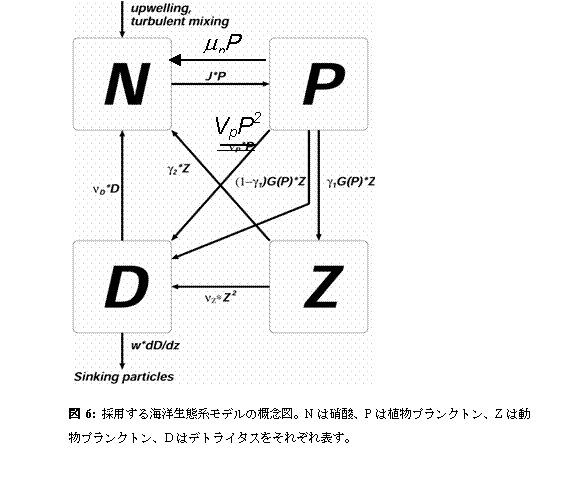
c.研究計画、方法、スケジュール統合モデル海洋炭素循環コンポーネントに組み込む生態系モデルとしては、Oschlies and Garçon(1999) による植物プランクトン、硝酸、動物プランクトン、デトライタスの4コンパートメント表層生態系モデルにOschlies(2001)による改変を加えたものを採用する(図6)。
このモデルの大循環モデルへの組み込みは2年目までで終了している。さらに陸域炭素循環モデルを組み込んだ結合モデルを用いて二酸化炭素漸増実験を3年目までに行い、論文執筆に着手する。
3年目以降は、この結合モデルを用いた実験及び結果の解析を継続して行う一方、鉄の大気輸送の効果も考慮した最先端のモデルを構築していくことを考えている。鉄の効果を取り入れた海洋生態系モデルは既にいくつか開発されてきており(e.g.,Leonard et al.,1999;Archer and Johnson 2000; Moore et al,2002)、それらを参考に我々のモデルを作り変えていくのは十分可能であると考えられる。また鉄分の大気輸送に関しては、研究実施者の一人が開発したダスト輸送モデルが大気大循環モデルにすぐ組み入れられる形で既に存在する。これらを組み合わせることで大気による鉄分輸送が生物ポンプに与える影響を陽に取り扱えるようになり、氷期−間氷期サイクルや地球温暖化に関して提案されている鉄を介したフィードバック機構(Kumar et al.1995)に関しより具体的な議論ができるようになると期待される。
d.平成15年度研究計画大気海洋結合気候モデルMIROCに、海洋炭素循環モデルと陸域炭素循環モデルとを結合させる作業に今年度中に着手する。海洋コンポーネントについては、4成分の簡略な生態系モデルと海洋大循環モデルとの結合がほぼ完了している。陸域コンポーネントについては、地球フロンティア研究システムで開発中のSim-CYCLEの気候モデルへの組み込みに15年度早々に着手する。陸面プロセスモデルMATSIROとSim-CYCLEの統合も行う。また亜寒帯林については温暖化により植生分布が大幅に変化することが懸念されるため、Sim-CYCLE拡張へ向け亜寒帯林に特化した植生動態モデルの開発を始める。MIROCへ組み込む前に各コンポーネントモデル単体での性能の吟味は充分に行う必要がある。 e.平成15年度研究成果e.1.モデルの設定平成14年度で海洋大循環モデルCOCOへの海洋生態系・炭素循環モデルの組み込みを終え、今年度はいくつかの異なったフォーシングのもとモデルを走らせ性能チェックを行った。モデルがもっともらしい振る舞いを見せるのを確認した後、大気海洋結合モデルMIROCに炭素循環モデル組み込みに取り組んだ。現在のところ、陸域と海洋の炭素循環を含む炭素循環−気候変化結合モデルの原型が完成した状態にある。以下本節ではモデル性能チェックを主目的として行ったa)海面水温・塩分、風応力といった海表面モデル駆動力に、観測に基づくものを用いたときと結合モデル出力を用いたときとの比較、b)人為起源二酸化炭素海洋吸収量の将来予測実験の結果について述べる。aのような比較を行うのは、b.で大気海洋結合モデルの出力で海洋モデルを駆動することの妥当性を検証するためである。 生態系モデルには前述の通り Oschlies and Garçon(1999)のモデルに、Oschlies(2001)が改良を加えたものを用いる。この生態系モデルに、海洋炭素循環モデル相互比較プロジェクト(Ocean Carbon Cycle Model Intercomparison Project,OCMIP)の推奨するプロトコルに従って炭酸系化学過程を導入し、海洋中の炭素循環を記述する。この炭素循環モデルを搭載する海洋大循環モデルCOCO(Hasumi,2000)の水平解像度は東西・南北ともに1度、鉛直の層数は54である。モデルの詳細については、平成14年度の成果報告書と、特に生態系モデルについてはOschlies and Garçon(1999)による記述を参考にされたい。 a.の比較実験について、観測に基づく駆動データとしては、海洋モデル相互比較プロジェクト(Ocean Model Intercomparison Project,OMIP)に対応して配布されている月平均データを用いる。モデル出力に基づく駆動データは、CCSR/NIESの大気海洋結合モデル(水平解像度は大気T21、海洋約2.8度)を用いIPCC第3次報告書(IPCC,2001)に提出した結果を基に作成する。すなわち、実験開始の1851年から、温暖化の兆候が見え始める1910年までの60年間について平均をとった出力データを海洋モデル用の駆動力として用いる。大気中二酸化炭素濃度としては、産業革命以前の値280ppmを与える。以下、観測に基づく実験を実験A1、モデル出力に基づくものを実験A2と呼ぶことにする。 b.の二酸化炭素海洋吸収量の将来予測実験としては、駆動力に結合モデルによる温暖化予測実験結果の時系列を用いるもの(実験B1)と、毎年一定の季節変動を用いるもの(実験B2)との2通り行う。B1とB2の結果を比較することにより、温暖化による海洋環境の変化が海洋中の炭素循環にもらたす影響についての知見を得ることができる。B2の駆動力にはA2と同じものを用いる。B1の駆動力には、上で述べたCCSR/NIESモデルによる温暖化実験の結果を直接用いる。大気中二酸化炭素濃度シナリオについては、B1,B2ともに1990年までは観測値に基づいた時系列を与え、以後は毎年1%ずつ漸増すると仮定した。以上、行った実験のまとめを表1に掲げる。
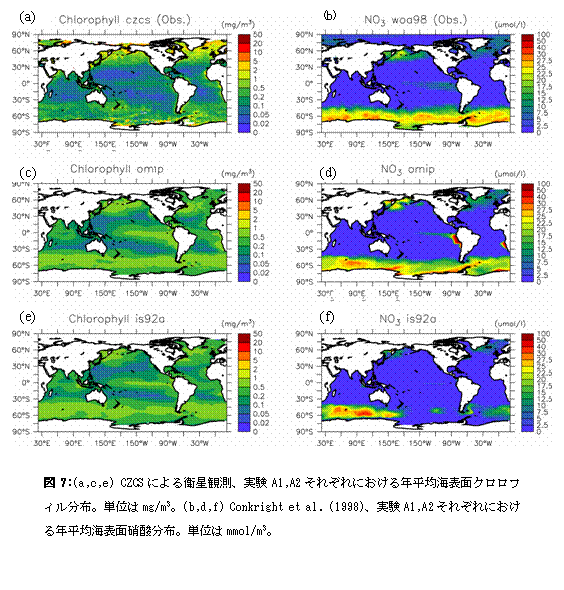
実験A1,A2の積分手順を以下に述べる。水温・塩分の初期値としては、Nakano(2000)が行った1万年の積分結果を、硝酸についてはConkright et al.(1994)による年平均気候値をそれぞれ与える。また全炭酸・アルカリ度については、海洋炭素循環モデル相互比較プロジェクト(Ocean Carbon-Cycle Model Intercomparison Project,OCMIP)に対応して山中ら(Orr,2002参照)のモデルにより得られた定常値を初期値として用いる。生態系モデルの硝酸以外の変数については、一定値0.1mmol/m3を初期値として与えた。この状態からA1,A2それぞれの駆動力を用いて19年間積分を行い、最後の1年間の結果について解析を行う。なお19年という積分期間は完全な定常状態を達成するには短すぎるものであるが、積分開始直後の衝撃を取り除くためには充分な期間であり、得られる結果は性能チェックのための解析に充分値する。 B1,B2については、A2の積分結果を初期値とし、それぞれに対応する表面駆動力、大気中二酸化炭素濃度をあたえて1851年から2100年まで積分する。以下ではまず、A1,A2の比較を行い、結合モデルの結果を海洋単体モデルの駆動力として使うことがどの程度妥当なものであるかを検討する。その後B1,B2の結果について述べ、人為起源二酸化炭素海洋吸収量の将来予測および温暖化による海洋環境の変化が炭素循環に与える影響について議論する。 e.2.計算結果e.2.1.A1,A2計算結果 図7は、年平均の海表面クロロフィル・硝酸濃度について実験A1,A2の結果および観測値を示したものである。なおモデル結果からクロロフィル濃度を得るには一定の換算比1.59(mg/m3)/(mmol/m3)を使用した。
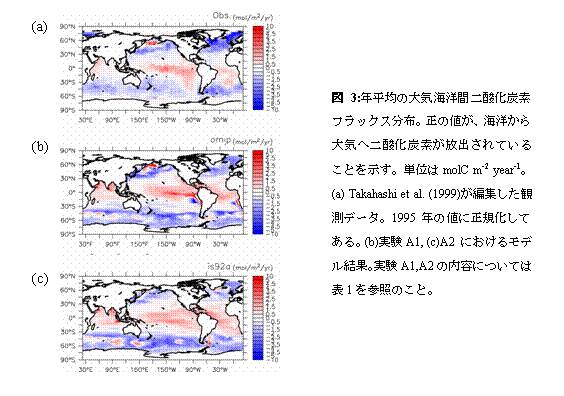
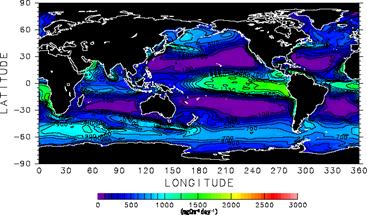
まずクロロフィル濃度について見ていくことにする。実験A1,A2とも、北太平洋・北大西洋北部の亜寒帯ジャイアや南極海、赤道域で高い値を示し、これは衛星観測に見られる特徴と一致している。ただし赤道域の極大については、とくに太平洋において両実験とも衛星観測に比べ高い値を示している。これは他の全球規模スケール海洋生態系モデルにも見られるバイアスであり(Fasham et al.2001)、その除去にはこれから時間をかけて取り組んでいく必要がある。 A1,A2両実験を比べると、クロロフィル濃度は値、分布ともよく似た結果を示していることが分かる。目に付く違いとしては、A1における太平洋赤道域の極大がA2のものより南北幅が大きいと言う点が挙げられる。この傾向は赤道域東部において特に顕著である。これは、この海域において赤道湧昇をもたらす貿易風が、実験A1に与えた風応力においてより強いことに起因する。また、後に述べるように実験A1ではペルー沖に強い沿岸湧昇が励起されており、このことも赤道域東部でこうした違いが見られる原因になっている。 次に硝酸濃度の分布について見ていく。Conkright et al.(1998)による気候値においては、北太平洋・北大西洋北部の亜寒帯ジャイアや南極海、赤道域で高い値が見られ、こうした特徴はA1,A2両実験においても大まかには再現されている。しかし実験A2においては、太平洋赤道域における硝酸極大の濃度は過小評価になっている。なおカラースケールの設定上A2では太平洋赤道域の硝酸極大が見えなくなっているが、クロロフィル濃度分布(図2e)に反映されているように実際には弱い極大が存在する。また実験A1では、ペルー沿岸域において硝酸濃度が顕著に高くなっている。これは、この海域での風応力の岸に沿う成分が、実験A1に与えたものにおいてより大きくなっているため、強い沿岸湧昇が励起されていることに起因する。実際、この海域における沿岸湧昇速度の典型的な値は1m/dayに達する。ここで採用しているような粗い格子サイズのモデルでは沿岸湧昇速度の最大値は過小評価される傾向にあり、赤道湧昇にも匹敵する1m/dayという湧昇速度は非常に大きいものと言える。Maier-Reimer(1993)が太平洋赤道域東部における非現実的な亜表層リン酸極大の形成要因として指摘した「栄養塩捕捉(nutrient trapping)」が、沿岸湧昇域でも発生したのがここでの非現実的な硝酸極大の形成要因である。観測値にもとづいたA1の駆動力の方が、ここで述べた点については非現実的な計算結果をもたらしている。 他に両実験結果間での違いのうち指摘すべきなのは、南極海の太平洋セクターにおける硝酸濃度であろう。A2におけるものの方が顕著に小さくなっている。これは南極海の冬季混合層が、A2で浅くなっているためである。結合モデル出力に基づいたA2の駆動力で冬季混合層が浅くなるのは、この海域でモデルの降水が客観解析データに比べ多くなっており海表面塩分が低く抑えられ、対流が起こりにくくなっていることによる。なおこの海域での海表面硝酸濃度にはエクマン湧昇流も大きな影響を及ぼすと考えられる。しかしながら今回のケースでは、風応力から計算されたエクマン湧昇流は実験A2においてむしろ強くなっており、これはA2の海表面硝酸濃度が低いことと矛盾する。エクマン湧昇流の違いが図2d,fの南極海表面硝酸濃度の違いをもたらしたのではないことが分かる。 図8には、大気海洋間の二酸化炭素交換量について、Takahashi et al.(1999)によってまとめられた観測データ、実験A1,A2それぞれの結果を示した。ただし、Takahashi et al.(1999)による分布は1995年現在に正規化されたものであり、大気中二酸化炭素濃度として産業革命以前の値280ppmを与えて得られたA1,A2の結果と直接比較することはできない。図8aを掲げたのは、あくまでおおよその分布・値の参考のためと捉えるべきである。
図8b,cを比較すると、太平洋赤道域でA1の方が湧昇が強いことに対応して図8bにおいて赤道に沿った二酸化炭素放出が図8cにおけるものより強くなっていることなどの違いが見られる。しかしながら、図7における実験A1,A2の比較から得られる印象よりは、図8b,cの結果はお互いに似ていると言えるであろう。大気海洋間の二酸化炭素交換量を決める重要な要素のひとつとして、全炭酸濃度がある。その全炭酸濃度と硝酸濃度はレッドフィールド比を通じてよく似た動態をもっているにも関わらずこうした違いが生ずるのは、大気海洋二酸化炭素交換そのものを通じたフィードバックが働くためである。南極海を例にとって説明する。図7fのように冬季混合層が浅く表層に硝酸が運ばれてこないときは、混合層以深からの全炭酸輸送も少なくなる。その結果、海表面の二酸化炭素分圧も低くなるが、すると大気との二酸化炭素分圧差が大きくなり、大気から海洋へ向かう二酸化炭素フラックスが増える。こうして表層付近の全炭酸濃度ひいては二酸化炭素分圧が増加し、結果として図8b,cの間で南極海の二酸化炭素交換量はさほど変わらなくなる。このようなフィードバック機構が存在するため、大気海洋間の二酸化炭素交換が図8b,cの間で似た分布を示しても、混合層付近の全炭酸の収支は大きく違っている可能性がある。 以上が、結合モデルの出力結果を用いて海洋モデルを駆動することの妥当性を検証するために行った比較研究の内容である。次節で述べる人為起源二酸化炭素海洋吸収量の将来予測を行う際に最も重要なのは海表面における二酸化炭素フラックスであるので、その意味では図8b,cにおけるA1,A2両実験の結果がよく似ていることは好ましい結果であると言える。ただし上で述べたように、海域によっては表層付近に全炭酸の収支が両実験で大きく異なっている可能性があり、二酸化炭素吸収量将来予測実験の解析ではその点に留意する必要がある。e.2.2.B1,B2計算結果 e.1.節で述べた手順に従い行った実験B1,B2について、全球積分した海洋二酸化炭素吸収量の時系列を図9に示す。実験B1において結合モデル内で作られる気候の年々変動に対応して二酸化炭素吸収量にも年々変動が見られることのほかは、両実験の結果は非常に良く似ている。2100年時点でのフラックスはB1,B2とも5PgC/year余りであり、両実験間での差は0.5PgC/year程度である。
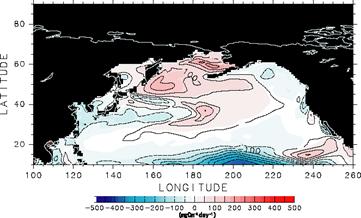
ここから分かるように、本モデルを用いた実験では、温暖化による海洋循環変化が二酸化炭素吸収量に与える影響は小さい。IPCC第3次報告書でも複数のモデルで同様な実験を行った結果が報告されており、それによると温暖化の影響はモデルによって差がある。しかし陸域炭素循環モデルで対応する実験を行った結果と比べると、どの海洋モデルでも温暖化の海洋炭素循環への影響は有意に小さくなっており、この点で我々の結果は過去のものと矛盾しないといえる。本プロジェクトで採用しているモデルはIPCC第3次報告書で採用されているものよりも海洋表層生態系の記述が詳細になっているが、ここでの温暖化実験では過去の実験結果を追認するに終わった。このこと自体は耳目を引く結果ではないが、陸域炭素循環と結合させる前段階として、我々のモデルがもっともらしい振る舞いを見せることは確認できたと言える。 ただし、IPCC第3次報告書に掲載されているモデル結果と比較すると、本モデルでの結果は温暖化の影響が小さい部類に属する。すなわち直感的には、実験B1では海洋が暖まるため海洋中の二酸化炭素分圧が高くなり、その結果B2よりも二酸化炭素吸収量が減ると考えられる。実際図9でもB1の方が吸収量は小さくなっているわけであるが、その差が他のIPCC第3次報告書のモデルより小さい傾向にある理由としては、次のような点が考えられる: 1)IPCC第3次報告書のモデルでは、温暖化の影響を評価する際、結合モデルに直接海洋炭素循環モデルを組み込んで計算を行っている。ここでの計算のように、海面水温などの海表面駆動力のみを取り替えて温暖化の効果を表現しようとすると、海洋内部での温暖化を再現しきれない可能性がある:2)温暖化の効果を入れた実験B1では、成層が強くなるせいで赤道湧昇が弱くなる。そのため高い全炭酸濃度を持った深層の水塊が表層付近に運ばれなくなり、赤道からの二酸化炭素放出が少なくなる。これは温暖化による海洋からの二酸化炭素放出を抑える方向に働く。ここで用いているモデルはIPCC第3次報告書のものより水平解像度が高く、赤道湧昇がより現実的に再現されるため、ここで述べたフィードバック機構がより効果的に働いた可能性がある: 3)水温が高くなって海洋中の二酸化炭素分圧が高くなっても、深層からの全炭酸供給が無ければ、大気への二酸化炭素放出によりすぐに二酸化炭素分圧はもとに戻ってしまう。深層からの全炭酸供給には鉛直拡散が大きな役割を果たすが、ここで用いたモデルはIPCC第3次報告書での多くのモデルよりも小さな鉛直拡散係数を採用している。そのため深層からの全炭酸供給が少なくなり、水温が高くなった効果が見えにくくなった可能性がある。 e.3.大気海洋結合炭素循環モデル開発状況 前節の結果を受け、大気海洋結合モデルへの炭素循環モデル移植にも着手し、現在その原型が完成した段階にある。結合モデルの分解能は、大気がT42L20、海洋が水平約0.5度(赤道付近)−1.4度(中高緯度)・鉛直44層である。当座のモデル開発のために用意した恣意的な設定と初期値を用い行った積分の3年目における陸面と海面からの年間二酸化炭素フラックス分布を図10に示す。また積分を行った3年間の地・海表面における全球平均二酸化炭素濃度の時系列を図11に示す。初期値が恣意的なものであるため図11には大きなトレンドが見られるが、トレンドを差し引いた季節変動振幅は4ppm程度と現実的な値を示している。ただし、図10,11はモデル開発状況について大まかな認識を読者に与えるためのものであり、科学的な知見を伝えるためのものではない。 今後、モデルパラメータのチューニングやコードの整備、充分なスピンアップを行い、炭素循環−気候結合モデル相互比較プロジェクト(C4MIP)への参加、ひいてはIPCC第4次報告書への貢献に備える。
簡単な生態系モデルNPZDとともに、動植物プランクトンをいくつかの機能別グループに分けて表現したモデルによる実験も行った。例えば、現在定数を用いている炭酸カルシウムの生成比をこのモデルの解析結果に基づいて、メカニズムに沿った経験的な関数を導入や、鉄制限などの導入など、NPZDの将来的な発展のいわば先導的な役割として反映させることができると思われる。このモデル(Global COCONUTS)は、東大気候センターで開発された海洋モデルCOCOの解像度1°x1°の全球モデルに、北太平洋の海洋科学機関(North Pacific Marine Science Organization:PICES)の低次生産モデルワークショップ(Lower Trophic Level Modeling Workshop)で開発された海洋生態系モデルNEMURO (North pacific Ecosystem Model Used for Regional Oceanography,)に炭酸系を拡張した海洋生態系-物質循環過程を組み込んだものである。このモデルは、世界中で開発されているもしくは開発中の生態系モデルと似た仕様を持っているが、主要な動物プランクトンであるカイアシ類の鉛直移住を表現しているところが特徴である。 Aita et al.(2003)では、カイアシ類の季節鉛直移住について議論を行った。この鉛直移住は、珪藻類が優占グループである西北太平洋や南大洋での基礎生産に影響を与え、カイアシが一年中表層にとどまるときには珪藻類による基礎生産を減少させる(図12)。一方、珪藻以外の植物プランクトンが優占している海域では、季節鉛直移住が行われない場合には、基礎生産が高くなる。これは、カイアシ類が、珪藻以外の植物プランクトンを捕食する小型動物プランクトンを捕食によって抑制するからである。カイアシ類の季節移住による炭素の下方輸送の全球総量は年間0.1ギガトンで、これは深さ1000mにおける沈降粒子による下方輸送量の約5〜10%に相当する。従って、海洋物質循環のより現実的な再現には、セジメントトラップで観測されるような沈降粒子による下層輸送に加え季節移住も将来的には必要となるだろう。
本年度は、1948年から2002年までの経年変動再現実験も行った。これは、NCEP再解析データセットとして提供されている海上気温・風応力・日射量・降水量・蒸発量などの日平均値を外力として、Global COCONUTSを駆動する計算である。全時間積分期間54年間を平均した生物基礎生産は、北太平洋亜寒帯域や南大洋、赤道などで高い値、亜熱帯域で低い値などをとるという大まかにいって観測で得られている分布を再現している(図13)。しかしながら、現バージョンでは、鉄制限などを考慮していないため、南大洋や赤道太平洋で過大評価である。これは速やかに改善すべき点である。
f.考察f.1.統合モデル開発の進捗について「d.平成14年度研究計画」に記載した事項は全て遂行できたと言える。C4MIPについても、大気海洋結合炭素循環モデルの結果提出期限は2005年5月であり、現在のペースで開発を進めていけば参加は充分可能である。C4MIPは国際的なプロジェクトであり全球炭素循環モデリングを行っている研究機関が多く参加している。また各種の期日設定等はIPCCへの貢献を意識しながら行っている。このプロジェクトへの参加が、共生プロジェクトの大きな目標の一つであるIPCC第4次報告書への貢献につながる可能性は高い。 f.2.NEMUROを用いた実験結果について 経年変動の再現において、数年スケールのENSO(El Nino and Southern Oscillation)や数十年スケールのPDO (Pacific Decadal Oscillation)などの気候変動に伴って、どのように生態系が応答するかを解析することによって、モデルの再現性、および、生態系内の変動がどのよう起こるのか(Plankton Dynamics)の理解を行うことができる。本年度はまだ解析が進んでいないが、1950年から2002年までのPDOと北太平洋中西部における生物基礎生産の偏差の関係について見てみた(図14)。PDOのインデックスはよく知られているように、気候ジャンプと呼ばれる1970年代に負の値から正の値へ変化している。それにほぼ対応して、北太平洋中西部における生物基礎生産は1970年代中期以降に減少しており、これは、Sugimoto et al.(1998)で述べられている結果と一致する。
g.参考文献Aita, M. N., Y. Yamanaka and M. J. Kishi (2003): Effect of ontogenetic vertical migration of zooplankton on annual primary production –Using NEMURO embedded in General Circulation Model-. Fish. Oceanogr., 12, 284-290.Archer, D. E., and K. Johnson, A model of the iron cycle in the ocean, Global Biogeochem. Cycles, 14, 1436-1446, 2000. Chai, F., M. Jiang, R. T. Barber, R. C. Dugdale and Y. Chao (2003): Interdecadal variation of the transition zone chlorophyll front: A physical-biological model simulation between 1960 and 1990. J. Oceanogr., 59, 461-475. Conkright, M. E., T. D. O’Brien, S. Levitus, T. P. Boyer, C. Stephens, and J. I. Antonov, World Ocean Atlas 1998, NODC, NOAA Atlas NESDIS 12, 1998. Conkright, M. E., S. Levitus, T. P. Boyer, NOAA Atlas NESDIS 1: World Ocean Atlas 1994, vol.1: Nutrients, 1994. Cox, P. M., R. A. Betts, C. D. Jones, S. A. Spall, and I. J. Totterdell, Acceleration of global warming due to carbon cycle feedbacks in a coupled climate model, Nature, 408, 184-197, 2000. Eslinger, D. L., M. B. Kashiwai, M. J. Kishi, B. A. Megrey, D. M. Ware and F. E. Werner (2000): Model task team workshop report. PICES Scientific Rep., 15, 1-77. Fasham, M. J. R. (Ed.), Ocean biogeochemistry: The role of the ocean carbon cycle in global change, IGBP Global Change Series, Springer-Verlag, Berlin Heiderberg New York, 336pp., 2003. Fasham, M. J. R., B. M. Balino, and M. C. Bowles, A New vision of ocean biogeochemistry after a decade of the Joint Global Ocean Flux Study (JGOFS), Ambio Special Report, 10, 4-30, 2001. Fridlingstein, P., L. Bopp, P. Ciais, J.-L. Dufrene, L. Fairhead, H. Letreut, P. Monfray, and J. Orr, Positive feedback between future climate change and the carbon cycle, Geophys. Res. Let., 28, 1543-1546, 2001. Hasumi, A., CCSR Ocean Component Model (COCO), CCSR Rep. 13, 68pp., 2000. IPCC, Climate Change 2001: The scientific basis, Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change, Houghton J. T. et al. (eds.), Intergovernmental Panel on Climate Change, Cambridge University Press, Cambridge New York, 2001. Kumar, N., R. F. Anderson, R. A. Mortlock, P. N. Froelich, P. Kubik, B. Bittrich-Hannen, and M. Suter, Increased biological productivity and export production in the glacial Southern Ocean, Nature, 378, 675-680, 1995. Leonard, C. L., C. R. McClain, R. Murtugudde, E. E. Hofmann, and L. W. Harding, An iron-based ecosystem model of the central equatorial Pacific, J. Geophys. Res., 104, 1325-1341, 1999. Maier-Reimer, E., Geochemical Cycles in an Ocean General Circulation Model: Preindustrial Tracer Distributions, Global Biogeochem. Cycles, 7, 645-677, 1993. Moore, J. K., S. C. Doney, D. M. Glover, and I. Y. Fung, Iron cycling and nutrient limitation patterns in surface waters of the World Ocean, Deep-Sea Res. II, 49, 463-507, 2002. Nakano, H., Modeling global abyssal circulation by incorporating bottom boundary layer parameterization, CCSR Rep., 14, 110pp., 2000. Orr, J. C., (ed.), Global Ocean Storage of Anthropogenic Carbon (GOSAC) Final Report, EC Environment and Climate Programme (Contract ENV4-CT97-0495), 2002. Oschlies, A., Model-derived estimates of new production: New results point towards lower values, Deep-Sea Res., 48, 2173-2197, 2001. Oschlies, A., V. Garçon, An eddy-permitting coupled physical-biological model of the North Atlantic 1. Sensitivity to advection numerics and mixed layer physics, Global Biogeochem. Cycles, 13, 135-160, 1999. Sugimoto, T., K. Tadokoro (1998): Interdecadal variations of plankton biomass and physical environment in the subarctic North Pacific. In: Biotic impacts of extratropical cliate variability in the Pacific. G. Holloway, P. Muller and D. Henderson (eds) Honolulu: SOEST Special Publication, pp51-60. Takahashi, T., R. T. Wanninkhof, R. A. Feely, R. F. Weiss, D. W. Chipman, N. R. Bates, J. Olafsson, C. L. Sabine, and C. S. Sutherland, Net sea-air CO2 flux over the ocean: An improved estimate based on air-sea pCO2 difference, In: Proc. 2nd Symposium on CO2 in the oceans, Nojiri, Y. (ed.), Tsukuba, Japan, January 18-23, pp. 9-15, 1999. h. 成果の発表<学会発表>Aita, M. N., Y. Yamanaka and M. J. Kishi: On ontogenetic vertical migration of zooplankton in GCM. 3rd International Zooplankton Production Symposium: "The Role of Zooplankton in Global Ecosystem Dynamics: Comparative Studies from the World Oceans", Gijon, Spain, May 20-23, 2003. M. Kawamiya and T. Matsuno,“Development of an integrated earth system model on the Earth Simulator”, IUGG2003, Sapporo, July 2003. M. Kawamiya, C. Yoshikawa, M. Aita and T. Matsuno,“Development of an integrated earth system model on the Earth Simulator -- Preliminary results from the ocean carbon cycle component –“, International Workshop on Earth System Modelliing, Hamburg, September 2003. M. Kawamiya,“Overview of Earth System Modelling in Japan” UK-Japan Workshop on Earth System Modelling, Cambridge, October 2003 M. Kawamiya, C. Yoshikawa, M. Aita, and T. Matsuno,“Projection of ocean uptake of anthropogenic CO2 using an ocean carbon cycle model: Preliminary results from the oceanic component of the integrated earth system model at FRSGC”, UK-Japan Workshop on Earth System Modelling, Cambridge, October 2003. Sasai, Y., A. Ishida, Y. Yamanaka, M. N. Aita and M. J. Kishi: Marine ecosystem and chemical tracer studies using two OGCMs. Final JGOFS Open Science Conference: "A Sea of Change: JGOFS accomplishments and the Future of Ocean Biogeochemistry", Washington DC, U.S.A, May 5-8, 2003. Yamanaka, Y., M. N. Aita and M. J. Kishi: Effects of ontogenetic vertical migration of zooplankton on simulations using NEMURO embedded in a General Circulation Model. EGS - AGU - EUG Joint Assembly, Nice, France, April 6-11, 2003. <論文出版> Aita, M. N., Y. Yamanaka and M. J. Kishi (2003): Effect of ontogenetic vertical migration of zooplankton on annual primary production –Using NEMURO embedded in General Circulation Model-. Fish. Oceanogr., 12, 284-290. Kawamiya, M. and A. Oschlies, “Impact of intraseasonal variations in surface heat and momentum fluxes on the pelagic ecosystem of the Arabian Sea”, J. Geophys. Res., 109, doi:10.1029/2003JC002107, 2004. 河宮未知生,「数値生態系モデルによる北太平洋の低次生産機構に関する研究」,海の研究,13, 135-150, 2004. 次のページ(1.3 陸域炭素循環モデルにおける植生態移動予測モデルの構築) |