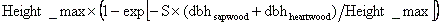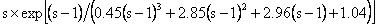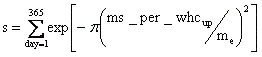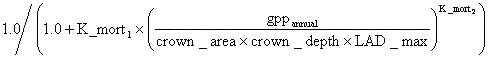1.炭素循環モデル、炭素循環・気候変化結合モデル3 研究結果の詳細報告へ戻る | HOMEへ戻る |
|||||||||||||||
1—3 陸域炭素循環モデルにおける植生帯移動予測モデルの構築担当機関:地球フロンティア研究システム
a.要約動的全球植生モデル(DGVM, Dynamic Global Vegetation Model)の設計、およびコード作成を進めた。このモデルの基本的なデザインは、陸域炭素循環モデルSim-CYCLEに、LPJ-DGVMの植生動態コンポーネントを組み合わせたものであるが、さらに林分の空間構造を明示的に組み込み、木本を個体ベースで扱うという拡張を行った。これらの拡張によって、森林ギャップの再生過程や樹木個体間の競争過程が的確に表現され、植生動態に伴う炭素収支や、気候変動に伴った植生分布変動の速度などを、これまで構築されてきたどのDGVMよりも正確に予測できることが期待される。現時点までに、一林分における計算を行うプログラムコードの開発がほぼ完了し、コードの最終チェックを行っている。今後、ベクトル化、並列化、パラメータ推定、調整等の過程を経て、平成16年度中までには全球グリッドでのシミュレーション結果を得る予定である。 b.研究目的気候環境は植生の構造と機能を強く規定するが、植生の構造と機能もまた、気候環境にフィードバック的な影響を与える(Foley et al. 2003)。このような過程を気候環境変動予測に取り込むために、共生第2プロジェクトで開発を進めている地球統合モデルKISSMEでは、陸域炭素循環モデルSim-CYCLE(Ito and Oikawa 2002)を一つの構成要素とする事を計画している。 Sim-CYCLEでは、植生の構造が水平方向に均質であることを仮定されており、しかも一度成立した植生は気候条件が大きく変動しない限り崩壊しない。しかし、発達した森林とは100m2前後の林分単位において崩壊と再生が繰り返される動的な構造体であり、このような動態(ギャップ動態)は、森林の炭素フラックス変動を強く規定している。なぜならば、一般に、成熟した林分では光合成生産量と維持呼吸量とがほぼ釣り合い、植生における炭素収支はプラスマイナス0近くで推移するが、他方、成長途中の林分では光合成生産物の多くが構造体の生産に用いられ、炭素のシンクとして機能するからである(例えば、Kira and Shidei 1967)。 また、Sim-CYCLEでは、植生タイプの分布が不変であることを仮定し、予め18種の植生タイプを陸面の各グリッドに割り振っている。しかし、百年から数百年といった長い時間スケールにおいては、植生の分布が気候条件の変動に伴って変化すると考えられ、したがってシミュレートする期間が長くなれば長くなるほど、モデル出力における潜在的な誤差が大きくなることが懸念されている。 そこで、そのような植生動態過程、および植生タイプ変化を地球統合モデルKISSMEで扱えるようにするため、Sim-CYCLEに死亡、攪乱、定着、競争といった植生動態過程を環境条件の関数として取り入れ、それらの結果に基づいて植生タイプの分布を変化させることで、いわゆる動的全球植生モデル(Dynamic Global Vegetation Model, DGVM)へと拡張する。 c.研究計画、方法、スケジュールc.1. 研究計画陸域炭素循環モデルSim-CYCLEへ以下の要素を追加し、動的全球植生モデルへと拡張する。 死亡、攪乱、定着といった植生動態過程を環境条件の関数として定義することによって、PFT間、または個体間の競争を組み込む。これらの動態過程のモデルは、基本的に、既存のDGVMであるLPJ-DGVM(Sitch et al. 2003)より得る。 さらに本モデルでは、木本を個体ベースで扱い、林分の空間構造を詳細に扱うという野心的な試みを行う。これによって、植生変化の方向と、それに要する期間の予測とがより適切に得られると期待される。なぜならば、植生変化とは、主として植物間の競争の結果として生じる現象であり、そしてそのような競争は局所的な光条件に強く規定されながら個体ベースで生じる現象だからである。 c.2. 方法(モデルの基本設計)この項において括弧内の記号は、大文字から始まるものについてはパラメーター、小文字から始まるものについては変数を示す。項の末尾に、それらの定義についての一覧表を添付する。またこれらはプログラム中のパラメーターや変数名とも対応している。 入力されるデータ、パラメーター(1)場所に関するパラメーター
出力される情報 (1)炭素動態
植物機能型と景観の求め方 全球グリッドにおいて植生動態を扱う上においては、一般的に陸上高等植物を少数の植物機能型(Plant functional types、以下PFTs)に要約させる。本DGVMではLPJ-DGVMの分類法に準じ、以下の10種類のPFTsを仮定した。PFT1~PFT8が木本、PFT9とPFT10が草本である。
各計算グリッドの植生景観は、優占する木本PFTの個体密度、及び優占する草本PFTの単位面積当たりバイオマスによって決定する。例えば、Tropical broad-leaved raingreenとC4草本とが優占している区画においては、木本の個体密度が高ければ「夏緑樹林」とし、木本の個体密度が低く草本バイオマスが高ければ「サバンナ」、木本の個体密度も草本バイオマスも低ければ「砂漠地帯」にするといった具合である。その分別基準はBIOME3(Haxeltine and Prentice 1996)より得る予定である。 木本の扱い 個体ベースで扱い、各個体は樹冠、幹、根の3部位から構成されるとした。各部位は次の変数で表現される。なお、樹冠と幹の形状は円柱を仮定し、根については形を明示的に扱わない。 樹冠:バイオマス(masscrown)、葉面積(la)、直径(crown_diameter)、深さ(crown_depth) 幹:バイオマス(masstrunc)、高さ(height)、地上1.3mにおける辺材と心材の直径(dbhsapwood,dbhheartwood) 根:バイオマス(massroot) 上記の変数に加えて、各個体は貯蔵資源(massstock)を有するとした。これは休眠期から展葉期に移る際に、葉の生産に用いられる資源である。 草本の扱い 草本は葉と根からのみ構成され、それぞれ単位面積あたりの重量(gmassleaf、gmassroot)によって表現される。また木本と同様に貯蔵資源(gmassstock)を持ち、休眠期から展葉期に移る際の葉の生産に用いられる。なお、計算区画においては、木本PFTと草本PFTとの分布域は重なることを仮定する。 定着(木本の場合) 木本は1m×1mのメッシュに1個体のみが生育できるとした。全ての新規定着木本は、胸高直径が0.01m(全て辺材部から構成される)とし、定着の際には、この胸高直径サイズを満たす木本の諸パラメーターを算出した。本モデルでは、樹冠が木本個体間で重ならないことを仮定する。したがって、新たに稚樹が定着できるメッシュとは、既存の樹木が生育していないだけではなく、稚樹が定着する際にその樹冠が既存個体の樹冠と重ならないことを条件となる。定着可能メッシュにおいては、昨年一年間の降水量が一定の閾値(Precmin)を超えている場合に、一定確率(P_establish)で木本が定着する。 定着可能なPFTを決定する基準については2つのパラメーターが与えられており、各グリッドの最近20年間の気候running meanがこれらの範囲に収まる場合に定着可能であるとした。(1)maximum coldest-month temperature(TCmax)、(2)minimum growth degree day (GDDmin)。これらの値はいずれもLPJ-DGVMより得た。 どの定着可能PFTがどのような割合で定着するのかについては、次の3つのシナリオから選択できる。実際のシミュレーションではScenario2と3を用いて、それぞれより植生変動速度の最大推定と最小推定を得る。もしこの両者の差が大きければ、それは植生変動における種子拡散の効果を無視できないことを意味する。 Scenario1(one specific PFT establish):予め指定した1種類のwoody PFTのみが定着する。パラメーター推定用。 Scenario2(infinite seed dispersal mode):現在どのようなwoody PFTが分布しているのかに関わらず、その環境条件で定着可能な全てのwoody PFTが等確率で定着する。 Scenario3(no seed dispersal mode):スピンアップにはScenario2を適用する。その後のシミュレーションでは、各woody PFT毎のバイオマスに比例して定着比率を決める。 定着(草本の場合) 草本は常に全グリッドに存在していると仮定し、定着過程を明示的に扱わない。草本は2種類のPFTから構成され、それぞれC3経路とC4経路を有するが、全ての草本はそのいずれの経路も利用する事ができると仮定し、前年のcoldest-month temperatureが15.5度(LPJの基準)以上ならばC4経路、それ未満の場合にはC3経路に切り替えられるとした。 水環境 水収支モジュールの大部分はSim-CYCLEと同一である。降水の一部は、林冠による遮断効果によって、直ぐに大気中に蒸発する。そして地上に届いた降水の一部または全部は、温度条件に応じて雪となり、これらは雪解けまで地表にストックされる。従って、地表面における水の入力は、降雨と雪解け水の2系統である。 土壌は地下0cm~30cmの上層(リター層)と、地下30cm~の下層(鉱物質層)の2層を仮定している。地表面に供給された降水は、土壌に与えられた浸透速度(Hyd_cond)に応じて、一部が上層から下層へと浸透し、最終的には次の4経路のいずれから外部へ抜ける:地表面からの蒸発(上層からのみ)、根からの吸い上げ(上層と下層から)、地表面からの乾燥(上層からのみ)、および流出(下層からのみ)。 上層と下層にはグリッド毎に最大含水量が与えられており(WHCup,WHClow)、実際の含水量をこの最大含水量で割った値を含水率(ms_per_whcup,ms_per_whclow)と定義した。各グリッドの水収支は、この含水率、プロット毎に予め与えられた浸透速度(Hyd_cond)、蒸散速度、そして各種気象条件の関数として決まる。 本モデルでは各PFTについて垂直方向の根のバイオマス分布をパラメーターP_root_upで定義する。そして、吸水が生じる土壌レイヤーは、このバイオマス分布に対応すると仮定した。すなわち、あるPFTの根バイオマスの80%が上層にあると定義した場合(i.e.,P_root_up=0.80)、このPFTにとっての土壌含水率は(water_status)は、0.8×ms_per_whcup+0.2×ms_per_whclowで与えられる。この計算の際、ms_per_whcupとms_per_whclowは、いずれも一週間または1ヶ月間のrunning meanを用いた。またPFT毎の根バイオマス垂直分布は、Jackson et. al. (1996)よりデータを得た。 光環境と光合成 植生に入る前の光合成有効放射量(par)は、Sim-CYCLEに準拠して、大気外側での短波放射量(radtop)と雲量(cloud)より、各グリッド毎に1日間隔で算出する。そのグリッド毎のparを用いて、各木本個体の樹冠へ入射されるparを算出する。まず全ての木本について、個体毎の開空度を算出した(月一回)。樹冠の頂上の中心全天に向かって放射状に256方向の直線を延ばし、その各線が通過する葉面積指数(lai)と通過する木本PFTに応じたlight attenuation coefficient(eK)からMonsi and Saeki(1953)の式に従って太陽光減衰率を求める。
草本は、光を巡る競争においては一方的に木本の抑圧下にあり、林床に届く光資源のみを利用できるとした。したがって、林分に照射された光の内、各木本の葉群で吸収された光量を差し引いた残りが、草本レイヤーに到達するとした。なお、草本レイヤー内の光減衰パターンについても、上のMonsi and Saeki (1953)のモデルを用いた。 以上のように求めた各木本個体の光条件、および草本レイヤーの光条件などを元にして、Sim-CYCLEに準拠して1日の積算光合成量を算出する。木本の光合成量は個体ベースで計算するが、このモデルでは木本個体間で樹冠が重ならないと仮定しているので、光合成量を樹冠ごとに積分する際には、その個体自身の葉群による自己被陰のみを考える。 呼吸とTurnover 維持呼吸と成長呼吸の2種類を扱う。基本的に、モデルとパラメーターは共にSim-CYCLEより得たが、Sim-CYCLEとは異なり、このモデルでは木本の芯材部は維持呼吸しないとし、また休眠期に入っている落葉性木本も維持呼吸しないとした。そして、利用可能な資源が維持呼吸に必要な必要な資源量に満たない場合には、まず貯蔵資源を用い(転換効率Efficiency_transout、転換ロスは大気中に放出)、それでも足りなければ木本・草本とも葉の重量の5%/日がリッターに組み入れられるとした。Turnoverはモデル、パラメーター共にLPJに準拠した。また、休眠期においてもturnoverのみは起きるとした。 分解 土壌炭素には短期間(数ヶ月から数年)で分解されるリター成分と、長期間(数十年から数世紀)で分解されるmineral soil成分があると仮定した。いずれの土壌炭素も、その分解速度は温度と土壌含水率の関数である。具体的な方法に関しては、Sim-CYCLEに準拠した。 展葉 各PFTsには、それぞれ落葉性か常緑性かの属性が与えられており、落葉性のPFTsにおいては、次の規則によって展葉期と落葉期が移り変わる。これらの規則は、パラメーターと共にLPJ-DGVMより得た。(※但し、このアルゴリズムだと、湿潤で温暖な環境下では、落葉性PFTであっても一年中葉をつけてしまうため、今後変更する可能性がある) 休眠期→活動期:・条件:最近一週間の平均気温がminimum base temperature(Tmpbase)を超え、なおかつ最近一週間の平均water_statusがminimum water stress factor(Ms_per_whcmin)を超える事。・イベント:貯蔵資源が利用可能となる。その際の転換効率はパラメーターEfficiency_transoutで与え(ここでは0.9を仮定)、転換ロスは二酸化炭素として大気中に放出されるとした。活動期に入ってからの展葉量は「最大展葉量×min[1.0,gdd/GDDreq]」で与え、gdd(growth degree day)の蓄積に従って徐々に上昇すると仮定した。ここでGDDreqは最大展葉までに必要とされるgddであり、これはPFT毎に定義されている。最大展葉量については、以下に算出方法を記載する。 活動期→休眠期:・条件:最近一週間の平均気温がminimum base temperature(Tmpbase)を下回る、または最近一週間の平均water_statusがminimum water stress factor(Ms_per_whcmin)を下回ること。・イベント:それまで展葉させていた葉が全てリッターに組み込まれる。なお、草本は全て多年生であることを仮定し、葉が落ちても根バイオマスおよびストック資源は残存すると考えた。 草本の最大展葉量:(1)根からの給水量による制約、(2)利用可能な資源量、の2種類の制約がかせられる。最大展葉量は、それぞれの制約が許す最大展葉量max1、max2のうちの最も小さな値である。
木本の最大展葉量:(1)樹冠サイズ、(2)辺材による水輸送量、(3)根からの給水量、(4)利用可能な資源量、の4種類の制約がかせられる。最大展葉量は、それぞれの制約が許す最大葉量max1,max2,max3、max4のうち最も小さな値である。
展葉ルーチンの後に成長ルーチンが毎日呼び出される。但し、休眠期間、および休眠解除から10日以内は呼び出しは生じない。成長ルーチンでは、次のプロセスが順に実行される。 (1)根の成長:まず過去一週間の平均林床parより、草本レイヤーにおける最適葉面積指数(laioptimum)を算出する。最適葉面積指数とは、草本葉群の最下部においてparが光補償点となる葉面積指数である。そして、この葉面積指数が達成された際に必要とされる根重を下式より求め、もし現在の根重がこれを下回れば、その不足分を(利用資源が無くならない限り)生産させる。
(2)Stock resource supplement:もし貯蔵資源量が、現在展葉している葉重を下回るようであれば、この両者が一致するまで貯蔵資源量を増大させる。資源の貯蔵に際しては、直接的なコストのみに着目しても、貯蔵場所からの転流、貯蔵化合物への転換、貯蔵細胞、組織、器官の構築などが必要とされる(Lambers et al., 1998)。そこで本モデルでは、分配された資源の一定割合(Efficiency_transin)のみが貯蔵資源となるとし、この際に生じるロスは大気中に二酸化炭素として放出されるとした。なお、Efficiency_transinには、0.9を仮定した。 (3)繁殖:この時点で利用可能な余剰資源は、全て繁殖に用いられると仮定し、リッターに組み入れた。 木本の成長 月に1度、成長ルーチンが呼び出される。但し落葉性woody PFTsについては、休眠期間、および休眠解除から30日以内は呼び出しは生じない。成長ルーチンでは、次のプロセスが順に実行される。 (1)樹冠の枯れ上げ:各PFTには、予めPAR補償点(パラメーター、PARmin)が与えられている。直前1ヶ月の平均PARが、このPAR補償点を下回る樹冠レイヤーがある場合、それを樹冠の最下部から順に10㎝刻みで枯れ上げさせ、crown_depthを調整する。なお、一度枯れ上げた樹冠レイヤーは、たとえ光環境が改善されたとしても、再び葉をつけることは出来ないとした。 (2)根の成長:最大展葉時において必要とされる根重を下式より求め、もし現在の根重がこれを下回れば、その不足分を(利用可能資源が無くならない限り)生産させる。
(4)幹の成長:この時点で残存している資源は幹の成長に用いられる。幹バイオマスは、胸高直径と木の高さから次式によって与えた。
但し、本モデルでは個体間で樹冠が同じ空間を占めない事を仮定するため、樹高の伸長に対しては空間的な制約が課せられる場合がある。そのような場合には、DBHがどれほど大きな値となっても、その制約以上には樹高は伸長できないとした。以上の制約の下で、そして残存している利用可能資源の範囲において実現可能な、最大sapwood幅成長量を数値的に求め、幹の幅と高さの成長を行わせる。 (5)樹冠断面積の増大:続いて、幹の成長量に応じて樹冠断面積を増大させる。樹冠径の最大可能値は、Reinecke's rule (Zeide, 1993)に基づき、次の関係で与えられる。
(6)繁殖:草本PFTと同じ。 攪乱 攪乱要因としては火災のみを考えた。火災モデルには、Thonicke et al. (2001)が開発し、Sitch et al. (2003)がLPJ-DGVMへの組み込みに際して簡略化したものを採用した。このモデルで火災は、fuel load(バイオマス+リッター)が200gC/m2以上蓄積している場合にのみ発生し、その場合の1年間の火災発生確率は、下式よりリッター含水率の関数として与えられる。
火災の生じた区画では、焼死した木本の全バイオマス、生き残った木本の葉群バイオマス、草本の全バイオマス、そして全てのリッターがCO2として放出されると仮定した。但し、一般的には火災跡には多くの炭化木片が残り、これらは生分解されにくいため炭素ストックとして長く地中に滞留する。また、それらの炭化木片が地上に散布されている状態ではアルベドが低下するとも考えられるが、本モデルはこれらの効果を考慮していない。火災時における木本の生存率には、PFT毎に特有の値が与えられる(パラメーター:Fire_resist)。この値もLPJ-DGVMより得たが、彼らはこれらの値の出所を引用していないので、どの程度信頼のおける数字であるかは不明である。 死亡(火災以外の要因による) 死亡は、木本PFTでのみ明示的に扱い、草本PFTでは高いTurnover rateによって暗示的に表現される。木本の死亡率は、Background mortality、Heat stress、Bioclimitic limit mortalityの合計であり、年に一度、個体毎に算出し、その値に応じて確率的に死亡させる。以下、各死亡率の定義について述べる。 Background mortalityは、その個体の置かれた光環境に応じて増減する確率的死亡であり、その確率は次式で与えられる。
Heat stressによる死亡率は、寒帯性木本PFTの3種(PFT No6,7,8)についてのみ下式に従って算出され、全体の死亡率に加算される。これは、モデル、パラメーター共にLPJ-DGVMより得た。
Bioclimitic limitによる死亡は、各PFT毎に定義された生存可能な環境範囲であり、モデル、パラメーター共にLPJより得た。その与え方は単純で、過去二十年間における最も寒い月の平均気温が、woody PFT毎に定められた値(パラメーター:TCmin)を下回れば、そのPFTは存続できないというものである。また、寒帯性落葉針葉樹(Boreal needle-leaved summergreen,PFT no.7)のみについては、最も寒い月と最も暖かい月との平均気温の差が、20年間の平均で43℃を下回った場合にも、存続できないとした。 シミュレーションの方法 T42(128×64)の各グリッドにおいて、30m×30m(※)の林分を10個独立にシミュレートさせ、その平均をもってグリッドの代表値とする。現バージョンではグリッド内の地理的ヘテロ性は扱わないので、この10個の林分が有するパラメーターは同一である。小さな林分を複数シミュレートさせる主な理由としては、攪乱の問題があげられる。例えば寒帯林で頻発する森林火災は、一度生じると、シミュレートしている林分の大きさに関わらず、その殆ど全てが壊滅してしまう。このように機会的に大きく変動する単一の林分から、グリッドの代表値を得る事は適当ではない。 :熱帯多雨林のように平均樹冠サイズの大きな植生が生じる地域については、100m×100m程度の大きさまで拡張する事も検討中である。 パラメーターリスト グリッドの位置情報
Altitude : altitude (m) 土壌パラメーター(グリッド毎に定義)
WHClow : water holding capacity below 30cm-depth soil (mm) Hyd_cond : soil hydraulic conductivity from upper to lower layer (day-1) Albedosoil : albedo on soil surface witout vegetation 形態、アロケーション(PFT毎に定義)
CA_max : maximum crown area (m2) LAD_max : maximum leaf area density (m2/m3) S : initial value of relative growth rate of height to diameter (m/m) Age_leaf : leaf age (year) SLA : specific leaf area (m2/g) FR_max : maximum leaf-to-root mass ratio (0.0-1.0) P_root_up : proportion of root mass within upper soil layer (0.0-1.0) Allometry1, 2, 3 : allometry parameter 1, 2, 3 (no dimension) 呼吸(PFT毎に定義)
SARGf, s, r : specific growth respiration rate for foliage, sapwood, root (g/g) 光合成(PFT毎に定義)
EK0 : light attenuation coefficient (no dimension) Lue0 : control light dependence coefficient (mol CO2 mol photon-1) Pmax : maximum stomatal conductance Tmpopt0 : optimum temperature for photosynthesis (Cecius) Tmpmin : minimum temperature for photosynthesis (Cecius) Tmpmax : maximum temperature for photosynthesis (Cecius) GSb0, b1, b2 : stomatal conductance parameter 1, 2, 3 (mmol H2O /m2/s) Km_nstl : maximum stomatal conductance Kmci : dependence of photosynthesis on intercellular CO2 concentration (ppmv) Cmpcd0 : CO2 compensation point (ppmv) その他の代謝(PFT毎に定義)
Efficiency_transin : energy transformation efficiency from available to stock resource Efficiency_trans out : energy transformation efficiency from stock to available resource フェノロジー(PFT毎に定義)
Status_waterbase : minimum base water status for foliation for dioceious PFTs (0.0 – 1.0) GDDreq : Growing Degree Day requirement to grow full leaf coverage Ms_per_whcmin : Minimum water stress factor for drought deciduous PFT 定着(PFT毎に定義)
Precmin : minimum precipitation for woody PFT establishment (mm/YEAR) TCmax : maximum coldest-month temperature (Cecius) GDDmin : Minimum growth-degree-day sum (5 Cedius degree base) 死亡(PFT毎に定義)
K_mort2 : parameter in background mortality equation (no dimension) TCmin : minimum coldest month temperature for survive (Cecius) Fire_resist : probability of survive when fire occurs (0.0-1.0) 乾燥重量
masstrunc : trunc mass of a woody individual (g in dyr-matter) massroot : root mass of a woody individual (g in dry-matter) massstock : stock mass of a woody individual (g in dry-matter) gmassleaf : leaf mass density of grass (g/m2 in dry matter) gmassroot : root mass density of grass (g/m2 in dry matter) gmassstock : stock mass density of grass (g/m2 in dry matter) 形態、形質
crown_diameter : crown diameter of a woody individual (m) crown_depth : crown depth of a woody individual (m) crown_area : cross section crown area (m2) dbhsapwood : sapwood diameter at 1.3m height (m) dbhheartwood : heartwood diameter at 1.3m height (m) la : leaf area of a individual (m2) laig : leaf area index of grass layer (m2/m2) 水環境
ms_per_whclow : moisture content per water-holding-capacity in lower soil layer (0.0-1.0) status_water : for each woody individual (0.0-1.0) 光環境
radtop : shortwave radiation at the atmosphere-top eK : light attenuation coefficient (no dimension) 気候
c.3.スケジュール平成16年度
5月 パラメーターの推定と調整用にデータを収集 6月-7月 パラメーターの推定と調整 8月 全球グリッドで運用できるようにコードを変更、並列化 9月 全球グリッドでのオフライン実験を地球シミュレーター上で行う 10月-翌3月 論文執筆、補足的なシミュレーション d.平成15年度研究計画Sim-CYCLEをベースとした動的全球植生モデルを設計、構築する。e.平成15年度研究成果これまでに、1グリッドにおける植生変動を扱うコードがほぼ完成し、今後、全球グリッドでの植生変動をシミュレートさせる下地を整えた。f.考察考察すべき結果は、まだ得ていない。g.引用文献Foley J. A., M. H. Costa, C. Delire, N. Ramankutty, and P. Snyder, Green surprise? How terrestrial ecosystems could affect earth's climate, Frontier Ecological Environment, 1, 38-44, 2003.Ito A., and T. Oikawa, A simulation model of the carbon cycle in land ecosystems (Sim-CYCLE): a description based on dry matter production theory and plot-scale validation, Ecological Modeling, 151, 143-176, 2002. Haxeltine A., and I. C. Prentice, BIOME3: An equilibrium terrestrial biosphere model based on ecophysiological constrains, resource availability, and competition among plant functional types, Global Biogeochemical Cycles, 10, 693-709, 1996. Haxeltine A., I. C. Prentice, and I. D. Cresswell, A coupled carbon and water flux model to predict vegetation structure, Journal of Vegetation Science, 7, 651-666, 1996. Hytteborn H., Deciduous woodland at Andersby, eastern Sweden, above-ground tree and shrub production, Acta Phytogeographica Suecica, 61, 1-61, 1975. Jackson R. B., J. Canadell, J. R. Ehleringer, H. A. Mooney, O. E. Sala, and E. D. Schulze, A global analysis of root distributions for terrestrial biomes, Oecologia, 108, 389-411, 1996. Kira T., and T. Shidei, Jap. J. Ecol., 17, 70-87, 1967. Lambers H., F. Stuart, I. Chapin, T. L. Pons, H. Lambers, and T. L. Pons, Plant Physiological Ecology, Springer Verlag, 1998. Leemans R., and I. C. Prentice, FORSKA, a general forest succession model, Meddelanden från Växtbiologiska Instotutionen, Uppsala (ISSN 0348-1417), 1989. Monsi M., and T. Saeki, Über den Lichtfaktor in den Pfanzengesellschaften und seine Bedeutung für die Stoffproduktion, Japapese Journal of Botany., 14, 22-52, 1953. Prentice I. C., W. Cramer, S. P. Harrison, et al., A global biome model based on plant physiology and dominance, soil properties and climate, Journal of Biogeography, 19, 117-134, 1992. Prentice I. C., and R. Leemans, Pattern and process and the dynamics of forest structure: a simulation approach, Journal of Ecology, 78, 340-355, 1990. Sitch S., B. Smith, I. C. Prentice, A. Arneth, A. Bondeau, W. Cramer, J. Kaplan, S. Levis, W. Lucht, M. Sykes, K. Thonicke, and S. Venevski, Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ Dynamic Vegetation Model, Global Change Biology, 9, 161-185, 2003. Thonicke K., S. Venevsky, S. Stich, et al., The role of fire disturbance for global vegetation dynamics: coupling fire into a Dynamic Global Vegetation Model, Global Ecology and Biogeography, 10, 661-677, 2001. Warning R. H., Estimating forest growth and efficiency in relation to canopy leaf aera, Advances in Ecological Research, 13, 327-354, 1983. Warning R. H., and W. H. Schlesinger, Forest Ecosystems: concepts and Management, Academic Press, Orlando, Florida, 1985. Zeide B., Primary unit of the tree crown, Ecology, 74, 1598-1602, 1993. h.成果の発表<口頭発表>発表者名:佐藤永、甲山隆司発表題名:Development of an integrated terrestrial ecosystem model for global changing prediction 発表場所等:種生物学会国際シンポジウム2003(2003年10月札幌) 次のページ(2.1 温暖化・大気組成変化相互作用(大気化学)) |
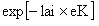



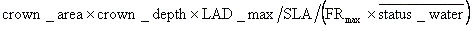
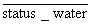 は、直前一週間ではなく、直前1ヶ月の平均値である。
は、直前一週間ではなく、直前1ヶ月の平均値である。