1.炭素循環モデル、炭素循環・気候変化結合モデル3 研究結果の詳細報告へ戻る | HOMEへ戻る |
||||||||||||||
1—3 陸域炭素循環モデルにおける植生帯移動予測モデルの構築担当機関: 地球環境フロンティア研究センター
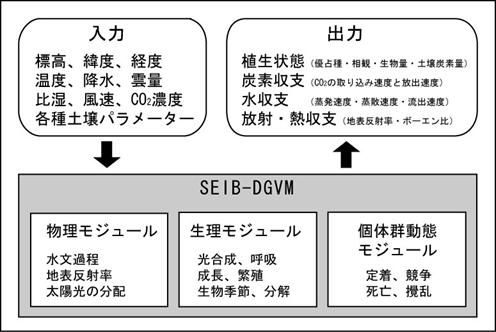
a.要約当グループは、プロジェクト期間中に、動的全球植生モデルSEIB-DGVMを完成させた。従来の同種のモデルに比較してSEIB-DGVMは、植物個体群動態を明示的に取り扱う点に特徴があり、これは植生変動予測の妥当性を高めるだけでなく、従来のモデルでは難しかった種類の感度分析を可能にする。その一例を示すために、全球植生分布の変化予測における種子分散力の影響を検討した。その結果、今後100〜200年の間に亜寒帯林の南限が北上するという予測が得られたが、その亜寒帯林が退いた広大な地域には、種子分散力を高く仮定した場合には温帯落葉樹林が速やかに進入し、種子分散力を低く仮定した場合には疎林帯に置き換わるという結果が得られた。また、この差に対応して、前者は後者に比べ全球バイオマス・全球NPPともに顕著に大きな値が得られた。この結果は、これまで全球シミュレーションで殆ど検討されることのなかった種子分散力の大きさが、植生の分布や機能を予測する上で、大きな影響を持つことを示すものである。 b.研究目的気候環境は植生の構造や機能を強く規定するが、植生の構造や機能もまた、蒸散・炭素循環・アルベドの変化などを通じて、気候環境にフィードバック的な影響を与える (Foley et al. 2003)。このような過程を気候環境の変動予測に含めるためには、生物地理化学過程や植生動態を取り込んだ陸域生態系モデルが必要とされる。その際に問題を複雑にしているのは、気候が変化しても、その新しい環境に適応した植物生態系が生じるまでに大きな時間遅れがあるという事である (Kohyama and Shigesada 1995, Kohyama 2005, Takenaka 2005)。このような時間遅れは、数十年〜数千年のオーダーで生じると推測されているが、これは種子の移入速度や既存植生の成長・更新頻度等々の関係で決まる複雑かつ多様な現象であり、いまだに信頼のおける予測は得られていない。そこで、現在地球が経験しているような、急速に進行する気候変化の元における植生帯の構造・分布・機能の過渡的変化をより的確にシミュレートするため、いわゆる動的全球植生モデル(DGVMs)が開発されてきた (reviewed by Cramer et al. 2001)。 当グループは、プロジェクト期間中に、動的全球植生モデルSEIB-DGVMを完成させた。従来のDGVMsと比較して、SEIB-DGVMを特徴づけているのは、グリッドボックスごとに幾つかの代表森林(または草地)をおき、その中に個体ベースで扱われた木本を定着させ、成長させ、そして死亡させる点である。定着した場所から移動することの出来ない植物にとって、例えば多少の気温上昇よりも、隣の木が枯れて光環境が改善される事の方が、よほど大きな環境変化であり、このような局所的に生じる個体間相互作用を無視しては過渡的な植生変化を的確に予測する事はできない、というのがこの設計を採用した理由である。 木本の定着を明示的に扱うSEIB-DGVMの特徴は、植生変化における種子分散の影響を評価することを可能にする。種子分散は、今後数百年間にわたって植生分布の変化を制御する重要な要素であると考えられているものの (Neilson et al 2005)、その効果は従来のDGVMでは検討されてこなかった。本サブテーマでは、SEIB-DGVMを用いて、次の100〜200年間における植生分布と植生機能の変化をシミュレートするが、このシミュレーションを様々な種子分散速度において実行することにより、種子分散が植生変化の予測に与える影響を評価する。 c.研究計画、方法、スケジュールモデルの構造について ここではSEIB-DGVMの概要を説明する。なお、モデルの詳細な構造はSato et al.(2007)に記載されている。SEIB-DGVMにおけるシミュレーションの基本単位は、空間構造を明示的に扱った30mx30mの仮想林分であり、この仮想林分に個々の木本が定着し、競争し、そして死亡する。他方、草本は単位面積あたりのバイオマスとして扱われ、木本の樹冠層の下に存在するとした。図19に、モデル全体の入出力と、SEIB-DGVMを構成する諸過程(物理・生理・動態の3つのモジュール群に分類されている)をまとめた。計算のタイムステップは3種類混在し、植物生理過程(土壌分解と幹の成長を除く)と物理過程は1日ごと、土壌分解と幹の成長は1ヶ月ごとで、植物動態過程と攪乱は1年ごとに計算される。人間による土地利用変化は、考慮していない。 SEIB-DGVMは、陸上維管束植物を、8種類の木本PFTと2種類の草本PFTによって扱っている。これらPFTの分類基準は、Sitch et al. (2003)から得た。これらのPFTは、単一の林分に共存可能である。木本PFTは個体ベースで扱い、各個体は樹冠・幹・根の3器官から構成されるとした。樹冠と幹の形状は円柱を仮定し、根については形態を明示的に扱っていない。各々の木本PFTには、定着可能な気候条件の範囲が予め定められている。
気候データについて シミュレーションに入力する気候データには、Cramer et al.(2001)のDGVM相互比較プロジェクトの為に用意されたGCM出力を使用した。このデータセットは、ハドレーセンターのHadCM2-SULモデルに、エアロゾルの観測値とIPCC-IS92aシナリオ(business-as-usual)のCO2濃度変化とを強制力として与えたものである(Houghton et al. 1992)。なお、この気候シミュレーションにおいては、生物化学過程から気候へのフィードバックは考慮していない。データセットは、上記のモデル比較プロジェクトのウェブサイトより入手し、SEIB-DGVMの計算時間間隔に適合するように、線形インバーションによって日ごとのデータに変換した。 シミュレーション条件 シミュレーションには、気候データに合わせて緯度2.5度×経度3.75度の格子系を使用し、各々の格子を30×30mの仮想林分一つによって代表させた。最初に、裸地より始まる500年間のスピンアップを実行した。このスピンアップでは、産業革命以前のCO2濃度(288ppm)と、そのCO2濃度の元における31年分のGCM出力を、繰り返し入力した。このスピンアップの結果を、産業革命以前の植生の状態と考え、本文の全てのシミュレーションにおける初期条件として使用した。 1861年から2099年へのシミュレーションを、次の3つの条件で実行した。
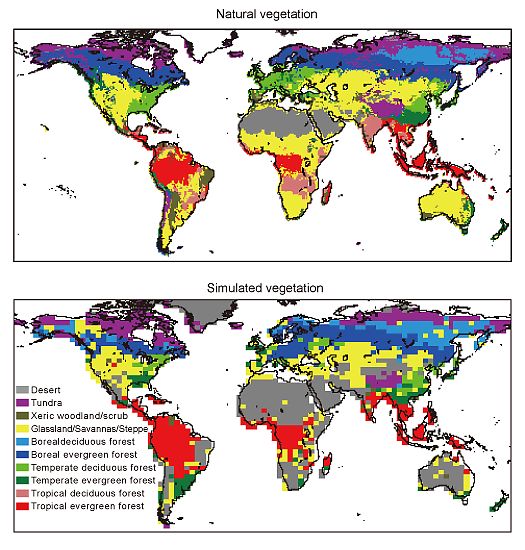
実験Cでは、IPCCのIS92amシナリオに従ってCO2濃度を変化させた。気候データには、スピンアップの際と同様に、産業革命前の条件におけるハドレーセンターGCMの出力31年分を繰り返し入力した。実験Tでは、CO2濃度は産業革命前の値を用い、気候条件には、IS92amシナリオにおけるハドレーセンターGCMの1861〜2100年のシミュレーション結果を入力した。実験CTでは、上の要領で、CO2濃度と気候条件の両方を変化させた。これらの環境変化に対する生態系応答のタイムラグを検討するために、全ての実験において、CO2を2100年で固定させた条件で、更に100年間のシミュレーションを行った。この期間、実験CTと実験Tでは、2100年のCO2濃度におけるGCM出力を入力した。 SEIB-DGVMでは、仮想林分の林床を1m×1mのセルに区切り、木本は個々のセルに1個体のみが生育できるとしている。また、樹冠が木本個体間で重ならないことを仮定しているので、新たに稚樹が定着できるセルとは、既存の樹木が生育していないだけではなく、稚樹が定着する際にその樹冠が既存個体の樹冠と重ならないことが条件となる。この条件に適合する定着可能なセルに対して、スピンアップの最初の50年間は、その環境条件で定着可能な全ての木本PFTを等確率で定着させた。そして50年目以降は、個々の木本PFTのバイオマスに比例させて、それぞれのPFTに定着可能なセル数を割り当てた。 スピンアップ後のシミュレーションの際には、定着可能なセルの10%、1%または0.1%を、その環境条件で定着可能な全ての木本PFTに均等に割り当て、残りのセルは、個々の木本PFTのバイオマスに比例させて、それぞれのPFTに割り当てた。上の定着条件を、それぞれ、種子分散シナリオrapid・mid・slowと名付けた。すなわち本実験では、3つの環境変動シナリオ(C・T・CT)と3つの種子分散シナリオ(rapid・mid・slow)の9種類の組み合わせにおいて、シミュレーションを行った。 d.平成18年度研究計画平成18年度は昨年度より引き続き、動的全球植生モデルSEIB-DGVMを改良し、現在の気候条件における植生の分布や機能の再現性力を高める。また、この改良されたモデルを用いて、オフラインによる温暖化実験を行う。 e.平成18年度研究成果産業革命以前における植生の分布と機能 図20は、500年間のスピンアップ後における植生分布地図を、自然植生地図と比較したものである。なお、これら分布図で用いた植生タイプの分別基準は、Haxeltine and Prentice (1996) を元に、Sato et al (2007) によって改変されたものである。また自然植生地図は、実際の植生分布データから作成されたものであり、気象データや生態モデルの出力に由来しない。SEIB-DGVMは、亜寒帯林帯と温帯林帯の分布をリーズナブルに再現した。最も顕著であった不一致は、乾燥地帯の植生帯に生じた。例えば、アフリカとインドの熱帯性雨緑林が広がる地域は、熱帯性多雨林帯と出力された。同様に、アフリカでは、乾燥地に適応した植生(草原/サバンナ/ステップ)の分布限界が、実際よりも湿潤地方に偏るという傾向が得られた。。
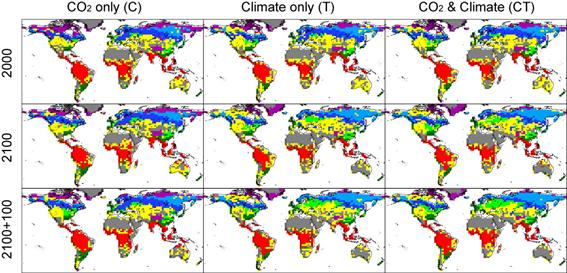
SEIB-DGVMによる全球の生態系機能の推定値は、他の観測やモデル研究で推定されている範囲に収まった。全球NPPと全球バイオマス炭素は、それぞれ44.7 Pg C / yr(スピンアップ最後の10年間の平均値)と585 Pg Cと推定された。他のモデルでは、それぞれの値を44.4〜66.3 Pg C / yr (Cramer et al. 1999) 及び557〜925 Pg C / yr (Sitch et al. 2003, Krinner et al. 2005, Kucharik et al. 2000, Foley 1995)と見積もっており、我々の値はこれらの下端に位置する。全球の土壌有機炭素量は1518 Pg C(1146Pg Cの土壌有機物+372Pg Cのリター)と推定され、この値は観測データを元にした推定値の範囲(1400〜1600Pg C, Schlesinger et al 2000)の中心に位置している。全球の流出水量は38.5×1015 kg / yr(スピンアップ最後の10年間の平均)と推定され、観測値を元にした推定値36×1015 kg / yr (Chahine 1992)に近い値が得られた。 環境条件を変化させた場合のシミュレーション結果 植生分布は気候変化に影響されたが、CO2濃度変化には殆ど影響を受けなかった。図21は、植生分布の変化パターンを、実験C・T・CT(種子分散シナリオは、いずれもrapid)間で比較したものである。いずれの実験においても、2000年までは顕著な変化は見られなかったが、それ以降に実験TとCTとで大きな変化が生じた。以下に主要なもののみ列挙する。 (1) まずユーラシア大陸において、温帯性落葉樹林と亜寒性落葉樹林(カラマツ帯)の北端がそれぞれ伸張した。またツンドラ地帯は、シベリアでは減少したが、ヒマラヤ高山帯やグリーンランドでは拡大した。これらの変化パターンは、実験TとCT間で殆ど違いは認められず、また実験Cでは殆ど植生分布に変化が生じなかったので、これらの植生分布変化は気候変化のみに起因していることが示された。
(2) 種子分散力を高く設定するほど、森林面積が拡大した。図22は、実験CTにおける植生分布変化パターンを、3つの種子分散シナリオ(rapid・mid・slow)の間で比較したものである。いずれの種子分散シナリオにおいても、ユーラシア大陸における亜寒性常緑樹林帯の南端が衰退したが、その後で置き換わる植生タイプは、種子分散シナリオによって異なった。すなわち、種子分散力の高いシナリオでは温帯性落葉樹林帯に、種子分散力の低いシナリオでは疎林帯へと変化した。このシナリオ間の差は、環境変化を停止させて100年経過した後にも残存しており、シミュレーションの最後まで植生変化が平衡に達しなかった事を示している。
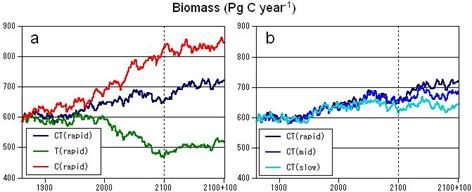
(3) CO2濃度と気候の変化は、生態系機能を逆方向に変化させた。図23a・24a・ 25aは、それぞれ全球におけるNPP・バイオマス炭素・流出水の経年変化を、実験C・T・CT(いずれも種子分散シナリオは"rapid")間で比較したものである。これら全ての項目において、実験Cは増加パターン、実験Tは減少パターン、そして実験CTは両者の中間パターンを示した。SEIB-DGVMでは、CO2濃度の増大は、光合成速度の増加、および気孔コンダクタンスの低下による蒸散量の減少をもたらすが、実験Cの結果はこれと対応している。また、入力した気候データで最も顕著な変化を示すのは平均気温の増大であるが、これは植物の呼吸速度を増加させ、光合成速度を乾燥域では減少させ亜寒帯域では増加させ、蒸発散量を増加させる。実験Tの結果は、これらの差し引きの結果と解釈できる。
(4) 環境変化実験におけるNPPと全球バイオマス炭素は、いずれも大きな種子分散力の元で高い値を持つ傾向が生じた。図23b・24b・25bは、それぞれ実験CTにおけるNPP・バイオマス炭素・全球流出水の経年変化を、異なる種子分散シナリオ間で比較したものである。NPPとバイオマスは、いずれも上昇傾向を示したが、その上昇幅は、高い種子分散力の元でより大きかった。これは、種子分散力が高いほど、森林面積が拡大したという結果、また新しい環境により適応した植生が速やかに広がり、より生産力を高くすると期待される点に対応している。これら種子分散シナリオ間の違いは、環境変化を停止させてから100年を経過した後でも残存しており、シミュレーションの最後まで植生変化が平衡に達しなかった事を示している。他方、流出水量においては、種子分散シナリオ間で顕著な差は認められなかった。 f.考察SEIB-DGVMは、産業革命以前における植生の分布と機能とを妥当な範囲で再現し、それらは環境変化に対して合理的な反応を示した。この環境変化に対する反応は、環境条件を固定して100年後においても持続し、それらが平衡に到達するまでは長い期間を要することが示された。このような植生の「慣性」は、種子分散に強く制限がかかる条件において、より強く働いた。例えば、実験CT(CO2濃度と気候条件の両方が変化する)において種子分散シナリオrapidとslowとの差を比較すると、NPPでは2100年で5.0 Pg C / Yr、2100+100年で7.3 Pg C / Yr、バイオマス炭素では36.3 Pg Cと77.5 Pg C であり、環境条件固定後の100年間においても種子分散シナリオ間の差は開き続けている。これらの結果は、今後100〜200年間における植生分布や植生機能の変化を予測する上において、種子分散が与える影響の大きさを示すものである。 なお、本実験でrapidと仮定している種子分散力では、定着可能なセルの10%を、与えられた環境条件で定着可能な木本PFTに等しく割り振っており、これは自然条件で実現可能な値とは思えない。したがって、これは人為的な種子散布や植林によって、植生変化を加速させた場合のシミュレーションと考えることができる。このように高い種子分散条件と現実的な種子分散力との間で、シミュレーション結果を比較することは、今後の植生変化に対して人間がいかに関与できるのか、提言を行うための基礎となるかもしれない。 今回のシミュレーションにおける種子分散の扱い方は、あくまでも一時しのぎ的なものに過ぎず、今後はより現実的な種子分散モデルを組み込むことが求められる。種子分散に関しては、既に様々なモデルが構築されている。風散布による種子分散を機構ベースで扱うモデルでは、単純な弾道方程式を利用することが一般的であり、母樹からの種子分散パターンを、種子が供給される高度・種子の落下速度分布・植生内の風速分布の関数として算出する (Greene and Johnson 1989)。このようなモデルは数百メートル範囲に散布される種子のみを対象とするが、風速変化の扱いを改善することで、上昇する気流に乗って遠距離まで散布される種子を扱うモデルも開発されている(Nathan et al 2002)。しかし、これら機構ベースの種子分散モデルが発達しても、それらを直接的に適用できるほどDGVMが扱う地理解像度は高くはならないだろう。そこでNeilson et al. (2002)は、景観スケールにおける種子分散パターンを、上のような機構ベースモデルによって高い地理解像度で再現し、その挙動をパラメーター化する方法を見いだすことで、DGVMに適用できる経験ベースモデルへとスケールアップさせることを提案している。 実際の種子分散には、地形・群落の断片化・種子を運搬する動物の増減・人間活動の影響など、様々な要素が関与し、地域ごとの特異性も高いはずなので、このような近似法は地域ごとに開発しなければならない。例えば、亜寒帯域における植生変化では、数十年から数百年周期で生じる森林火災が強く関与しており、森林火災後のみに定着できる樹種も多い(Purdy et al. 2002)。これは森林火災が、地表を厚く被っていた地衣やリターを取り除き、稚樹が定着できる土壌面を露出させるからである。このような過程は、地域や植生タイプごとの特異性が高いし、また機構ベースでモデル化するには複雑すぎる。したがって、特定の地域や植生ごとに得られている植生タイプ変化の経験則を、その地域・生態系のみで適用される強制力という形でモデルに取り込むこと事が現実的かもしれない。たとえば、アラスカの植生変化を再現することを目的としたシミュレーターALFRESCO (Starfield and Chapin 1996)では、攪乱や遷移に伴った植生タイプの変化パターンを、与えられた気候環境ごとの経験則を適用することで計算している。今後、生物地球化学モデルが進むべき方向のひとつは、全球モデルとしての枠組みを保ちながら、このような地域・生態系に特異的な事象を取り込んでいくことであろう。 g.引用文献Chahine MT (1992) The hydrological cycle and its influence on climate, Nature 359:373-380 Cramer W, Kicklighter DW, Bondeau A, Moore B, Churkina C, Nemry B, Ruimy A, Schloss A (1999) Comparing global models of terrestrial net primary productivity (NPP): overview and key results. Global Change Biol 5:1-15 Cramer W, Bondeau A, Woodward FI, Prentice IC, Betts RA, Brovkin V, Cox PM, Fisher V, Foley JA, Friend AD, Kucharik C, Lomas MR, Ramankutty N, Sitch S, Smith B, White A, Young-Molling C (2001) Global response of terrestrial ecosystem structure and function to CO2 and climate change: results from six dynamic global vegetation models. Global Change Biol 7:357-373 Foley JA, Costa MH, Delire C, Ramankutty N, Snyder P (2003) Green surprise? How terrestrial ecosystems could affect earth's climate. Front Ecol Environ 1:38-44 Greene DF, Johnson EA (1989) A Model of Wind Dispersal of Winged or Plumed Seeds. Ecology 70:339-347 Haxeltine A, Prentice IC (1996) BIOME3: An equilibrium terrestrial biosphere model based on ecophysiological constraints, resource availability, and competition among plant functional types. Global Biogeochem Cy 10:693-709 Houghton JT, Callander BA, Varney SK (eds) (1992). Climate Change 1992 - the Supplementary Report to the IPCC Scientific Assessment. Cambridge University Press, Cambridge. Kohyama T (2005) Scaling up from shifting-gap mosaic to geographic distribution in the modeling of forest dynamics. Ecol Res 20:305-312 Kohyama T, Shigesada N (1995) A size-distribution-based model of forest dynamics along latitudinal enviromental gradient. Vegetatio 121:117-126 Krinner G, Viovy N, de Noblet-Ducoudre N, Ogee J, Polcher J, Friedlingstein P, Ciais P, Sitch S, Prentice IC (2005) A dynamic global vegetation model for studies of the coupled atmosphere-biosphere system. Global Biogeochem Cy 19:GB1015 Kucharik CJ, Foley JA, Delire C, Fisher VA, Coe MT, Lenters JD, Young-Molling C, Ramankutty N, Norman JM, Gower ST (2000) Testing the performance of a Dynamic Global Ecosystem Model: Water balance, carbon balance, and vegetation structure. Global Biogeochem Cy 14:795-825 Nathan R, Katul GG, Horn HS, Thomas SM, Oren R, Avissar R, Pacala SW, Levin SA (2002) Mechanisms of long-distance dispersal of seeds by wind. NATURE 418:409-413 Neilson RP, Pitelka LF, Solomon AM, Nathan R, Midgley GF, Fragoso JMV, Lischke H, Thompson K (2005) Forecasting regional to global plant migration in response to climate change. BioScience 55:749-759Purdy BG, Macdonald SE, Dale MRT (2002) The regeneration niche of white following fi re in the mixedwood boreal forest. Silva Fennica 36:289-306 Sato H, Itoh A, Kohyama T (2007) SEIB-DGVM: A New Dynamic Global Vegetation Model using a Spatially Explicit Individual-Based Approach. Ecol Model 200: 279-307 Schlesinger WH, Winkler JP, Megonigal JP (2000) Soils and global carbon cycle. In: The Carbon Cycle (eds. T.M.L. Wigley & D.S. Schimel), pp93-101. Cambridge University Press, Cambridge. Sitch S, Smith B, Prentice IC, Arneth A, Bondeau A, Cramer W, Kaplan JO, Levis S, Lucht W, Sykes MT, Thonicke K, Venevski S (2003) Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model, Global Change Biol 9:161-185 Starfield AM, Chapin FS III (1996) Model of transient changes in arctic and boreal vegetation in response to climate and land use change. Ecol Appl 6:842-864 Takenaka A (2005) Local coexistence of tree species and the dynamics of global distribution pattern along an environmental gradient: a simulation study. Ecol Res 20:297-304 h.成果の発表<口頭発表> Sato H, An introduction of SEIB-DGVM, a new dynamic global vegetation model using a spatially explicit individual-based approach, and its applications to tropical rain forest, International Symposium on global change on vegetation and cycling of materials in ecosystems (札幌), 2007年2月 佐藤永, 伊藤明彦, 甲山隆司, 動的全球植生モデルSEIB-DGVMによる、植生の構造と機能の変動予測, 日本地球惑星科学連合2006年大会 (幕張), 2006年5月 佐藤永, 植生動態モデルにおけるデータ利用, 第54回日本生態学会大会(松山), 2007年3月 <論文出版> Sato H, Itoh A, Kohyama T, SEIB-DGVM: A New Dynamic Global Vegetation Model using a Spatially Explicit Individual-Based Approach, Ecological Modelling 200:279-307, 2007 佐藤永, 生物地球化学モデルの現状と未来 静的モデルから動的モデルへの展開, 日本生態学会誌(投稿中) 次のページ(2.1 温暖化・大気組成変化相互作用(大気化学)) |