プレスリリース
国立研究開発法人海洋研究開発機構
立命館大学
久留米大学
ゲラニルゲラニル還元酵素の新規な水素付加反応特性を発見
~酵素の産業利用や光合成色素の進化解明の糸口となる可能性~
1. 発表のポイント
- ◆
- ゲラニルゲラニル還元酵素とは、植物、微生物、古細菌などの広範な生物が持つイソプレノイド含有化合物(クロロフィル色素、ビタミンE、ビタミンK1、膜脂質等)の不飽和炭素結合に対する水素付加反応を触媒する酵素である。
- ◆
- 好塩性光合成細菌の一種であるHalorhodospira halochlorisが持つゲラニルゲラニル還元酵素が、4水素付加型の反応特性を持つことを明らかにした。これは今までに植物、藻類、シアノバクテリアを含むいずれの光合成生物でも知られていない特異な酵素反応特性である。
- ◆
- 本菌の酵素は、ゲラニルゲラニル還元酵素一般の作動原理を解明していく足掛かりとなり、今後は酵素の任意改変と産業利用や、光合成色素の進化研究の発展に役立つことが期待される。
2. 概要
国立研究開発法人海洋研究開発機構(理事長 松永 是)超先鋭研究開発部門 超先鋭研究プログラムの塚谷祐介副主任研究員は、立命館大学、久留米大学と共同で、好塩性光合成細菌の一種であるHalorhodospira halochlorisが持つゲラニルゲラニル還元酵素が新規の水素付加反応特性を持つことを明らかにしました。
植物や微生物が持つクロロフィル色素、ビタミンE、ビタミンK1、あるいは古細菌の膜脂質といった生体化合物には、イソプレノイドと呼ばれる疎水性(水に溶けにくい)部位(図1)があり、これは細胞膜への結合あるいは細胞膜そのものとしての機能にとって重要な部位です。ゲラニルゲラニル還元酵素はこうした生体化合物のイソプレノイド部位にある不飽和炭素結合への水素付加反応(還元反応)を担う酵素であり、この反応によってこれら生体化合物は正常な結合・機能を果たすことができます。特に光合成生物の色素合成経路においては、クロロフィル(葉緑素)あるいはバクテリオクロロフィルの最終合成反応を触媒することが古くから知られていました。しかしながら、ゲラニルゲラニル還元酵素がどのようなメカニズムによって反応が起きるのかという作動原理は依然として分かっていません。
今回の研究では、光合成細菌の一種であるHalorhodospira halochlorisのバクテリオクロロフィル色素合成系で働くゲラニルゲラニル還元酵素が、これまで知られてきた6つの水素原子を付加する反応ではなく、光合成生物として初めての4水素付加型の特異な酵素活性を持つことを発見しました(図2、図3)。さらに、本酵素のアミノ酸を一部変化させることで、これも初めてとなる特異な2水素付加型の活性を示す酵素へと改変することにも成功しました(図3)。
この水素付加反応は、クロロフィル色素合成の最終段階で行われるものですが、今回の研究では、いずれの酵素反応の結果生じる色素でも光合成ができることが確認されており、今後は異なる色素が生じた理由などの解明に取り組むことで、光合成進化の中で色素分子種が果たす役割などを明らかにしていきます。光合成反応の要となるクロロフィル類の生合成については、未だ解明されていないことが多く、本研究がクロロフィル合成の全容把握や光合成色素の進化研究の発展に繋がることが期待されます。
今回の研究によって、3つのタイプの水素付加反応を触媒する酵素が出揃ったことになり、この研究成果はゲラニルゲラニル還元酵素一般の作動原理を解明する足掛かりとなるものです。将来的にはビタミンやキノン等の健康食品化合物の改変・生産へ役立つことも期待されます。
なお、本研究の一部は科学研究費補助金(19H02018、18H03743、17H06436)およびアストロバイオロジーセンター プロジェクト研究(AB021015)によって実施されました。
本成果は、アメリカ微生物学会が発行するJournal of Bacteriology誌に3月15日付けで掲載されました。
- 国立研究開発法人海洋研究開発機構
- 久留米大学
- 立命館大学
3. 背景
植物や微生物のクロロフィル色素、ビタミンE、ビタミンK1、あるいは古細菌の細胞膜脂質といった生体化合物には、疎水性のイソプレノイド側鎖部位(図1)があり、この部位は細胞膜への結合あるいは細胞膜そのものとしての機能に必須です。これら化合物の生合成(生物がその構成成分である生体分子を作り出すこと)の過程において、イソプレノイド部位に存在する不飽和二重結合は、ゲラニルゲラニル還元酵素の働きによって水素原子を付加されて単結合となることが知られています(図2左:クロロフィル色素の場合を例示)。この水素付加反応によって、イソプレノイド含有化合物は正常な膜への結合・機能を果たすことができます。しかしながら、ゲラニルゲラニル還元酵素の作動原理は今でも分かっていません。
クロロフィル色素は、光を吸収するためのクロリン部位と、膜への結合に必要なイソプレノイド部位で構成されます(図1)。クロロフィル色素が生体内で合成されるためには、まずクロリン部位が多数の酵素による修飾反応を経て合成され、その後イソプレノイド部位がクロリン環の17位に結合され、最後にその結合したイソプレノイド部位の修飾反応、すなわちゲラニルゲラニル還元酵素による水素付加反応が起こります(図4)。
今回研究対象とした、光合成細菌の一種であるHalorhodospira halochlorisは、塩湖などに見られる好塩性の生物です。本研究でこの細菌に着目した理由は、光合成細菌で一般的に広くみられるバクテリオクロロフィル色素には不飽和二重結合が1つしかないイソプレノイド部位(これをフィチル基と言う)が付いていますが、Halorhodospira halochlorisの色素には不飽和結合が2つあるイソプレノイド部位(これをテトラヒドロゲラニルゲラニル基と言う)が付いていたからです(図2)。つまり仮説として、Halorhodospira halochlorisのゲラニルゲラニル還元酵素の水素付加反応性が一般的なものとは異なる可能性が予想されました(あるいは研究開始当初は本菌にはゲラニルゲラニル還元酵素がそもそも存在しない可能性もありました)。2019年に我々はHalorhodospira halochlorisの完全長ゲノム解読結果を論文発表し(参考文献1)、その研究によって本菌ゲノムにはゲラニルゲラニル還元酵素をコードする遺伝子が存在することを見出しました。そこで今回の研究では、同定したHalorhodospira halochlorisのゲラニルゲラニル還元酵素の反応特性を調べました。
4. 成果
今回の研究では、一般的なバクテリオクロロフィルを持つことが分かっているモデル光合成細菌Rhodobacter sphaeroidesを用いて、この細菌のゲラニルゲラニル還元酵素をコードする遺伝子を、Halorhodospira halochlorisの当該遺伝子と入れ替える実験を行いました。その結果、遺伝子を入れ替えたRhodobacter sphaeroidesの株は、フィチル基ではなくテトラヒドロゲラニルゲラニル基をもつバクテリオクロロフィルを生産するように変化しました。このことはつまり、導入したHalorhodospira halochlorisのゲラニルゲラニル還元酵素が水素付加の少ない特異な反応特性を持つためであることを直接的に示しています。具体的には、Halorhodospira halochlorisのゲラニルゲラニル還元酵素は、バクテリオクロロフィルのイソプレノイド部位に対して、通常の6つの水素を付加する反応ではなく、4水素付加型であることが分かりました。これは今までに植物、藻類、シアノバクテリアを含むいずれの光合成生物でも知られていない特異な酵素反応特性です。
また、Halorhodospira halochlorisのゲラニルゲラニル酵素の活性に重要な酵素部位を調べるために、酵素タンパク質を構成するアミノ酸のうちいくつかの領域を改変したゲラニルゲラニル酵素を作成したところ、元の酵素に比べてさらに水素付加数が少ない、2水素付加型の活性を示すように変化したものを見出しました。これは、改変したアミノ酸領域が、元酵素の活性の一部に重要な領域であることを示唆しています。
通常のゲラニルゲラニル還元酵素が本来備えている反応特性は、2つの水素を付加する還元反応を3回連続して行う(合計で6水素付加)という、単一の酵素が触媒する反応としては珍しいものです。そのような複雑な反応が、酵素内のどのような作動メカニズムによって起こっているのかはいまだに分かっていません。今回の研究によって、3つのタイプの反応を触媒するゲラニルゲラニル還元酵素(通常タイプ、Halorhodospira halochlorisタイプ、改変型Halorhodospira halochlorisタイプ)が出揃ったことになります。このことは、異なるタイプの酵素の比較解析を通じてゲラニルゲラニル還元酵素一般の作動原理を解き明かしていく上で極めて重要な成果です。
5. 今後の展望
ゲラニルゲラニル還元酵素は、クロロフィル類の生合成経路の最終ステップで機能する(図4)ことから、光合成進化の中では比較的最近に獲得されたものであると考えられます。色素合成酵素の進化的獲得経緯や生理的意義の解明が進むことによって、始原的な光合成機構や酵素が獲得された当時の地球環境推定といった地球と生命の共進化の研究発展へ繋がることが期待されます。
今回の研究では、クロロフィル色素生合成で働く酵素として、これまでに例のない反応形態が見つかりました。一方で、ゲラニルゲラニル還元酵素を入れ替えたモデル生物Rhodobacter sphaeroidesの変異株は、色素のイソプレノイド部位が変わったにもかかわらず、光合成で生育することが可能でした。また、Halorhodospira halochlorisが元々生産する色素についても、通常とは異なるイソプレノイド部位を持つことが、本菌の光合成反応にどのような影響をもたらしているのかまでは解明できていません。つまり酵素の反応性の違いが生物に何をもたらしているのかという根源的な問いは残されたままです。色素のイソプレノイド部位の二重結合数の変化は、吸収する光波長には影響せず、色素と細胞膜や膜タンパク質との相互作用に関係があると予想されますが、本菌が棲息する塩湖などの高塩濃度環境との関連性も含めて、その生物的意義を明らかにしていくことが今後の課題の一つです。 今回の研究成果により、通常の6水素付加型に加えて、Halorhodospira halochlorisの4水素付加型、さらにそれを改変した2水素付加型のゲラニルゲラニル還元酵素が見出されました。これら3タイプの酵素について、生化学的・構造学的な検証と比較を進めていくことで、ゲラニルゲラニル還元酵素の連続水素付加反応の作動原理を解明できる可能性が高まったと言えます。単一の酵素内で複数回の還元反応を行うという特性の普遍的原理を理解できれば、将来的には、ビタミンやキノン等のイソプレノイド化合物の任意改変と生産に役立つこと、さらには水素原子の出し入れによるエネルギーの貯蔵・輸送といった酵素の産業利用へと繋がることが期待されます。
【用語解説】
- ※1
- イソプレノイド:
炭素数5のイソプレン単位を基本骨格として持つ天然有機化合物の総称。例えばカロテノイド、ステロイド、キノン、ビタミンといった化合物の多くがイソプレノイドに含まれる。化合物の一部分がイソプレノイドから成ることもあり、例えばクロロフィル類の17位のゲラニルゲラニル側鎖やフィチル側鎖(図1、図2参照)はイソプレノイド構造をした部位である。
- ※2
- ゲラニルゲラニル還元酵素:
クロロフィルの生合成ステップにおいて、クロリン環の17位に結合したゲラニルゲラニル基(イソプレノイドの一種)をフィチル基へと変化させる反応を触媒する(図2左側を参照)。
- ※3
- 不飽和炭素結合:
炭化水素化合物における炭素―炭素間の二重結合(あるいは三重結合)を不飽和炭素結合と呼ぶ。
- ※4
- コードする遺伝子:
遺伝子がタンパク質をコードするとは、特定の遺伝子の核酸塩基配列にしたがって、特定のタンパク質がタンパク質合成で作られること。
【参考文献】
- 1.
- イソプレノイド:
Tsukatani, Y., Hirose, Y., Harada, J., Yonekawa, C., and Tamiaki, H. (2019) Unusual features in the photosynthetic machinery of Halorhodospira halochloris DSM 1059 revealed by complete genome sequencing. Photosynthesis Research, vol.140, pp.311−319.
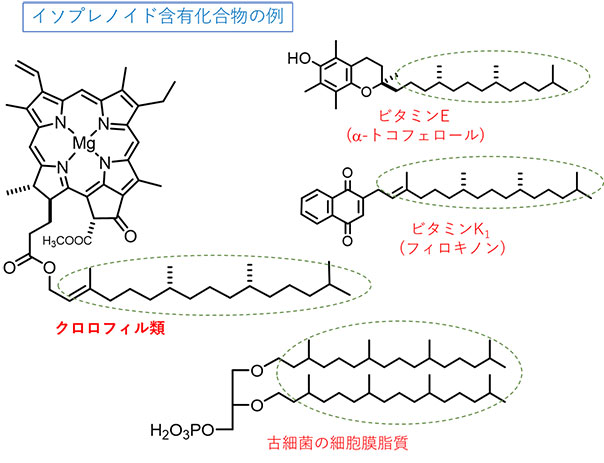
図1 イソプレノイド部位を持つ有機化合物の例
イソプレノイド化合物は自然界に多く存在し、ここには代表的な例としてクロロフィル類、ビタミン類、古細菌由来の膜脂質の化学構造を示す。点線の囲い部分がイソプレノイド側鎖部位である。
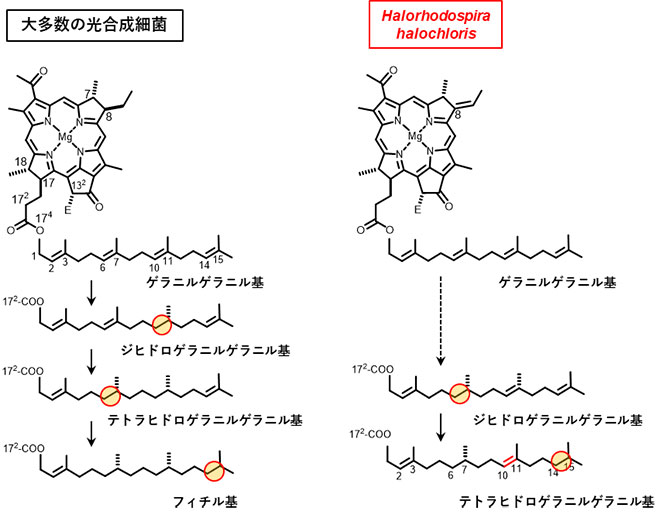
図2 クロロフィル色素類の生合成経路におけるイソプレノイド部位の還元反応
左:クロロフィル色素類17位に結合したイソプレノイド部位は、生合成の初めはゲラニルゲラニル基の状態である。ゲラニルゲラニル還元酵素による水素付加数に応じて、ゲラニルゲラニル基→ジヒドロゲラニルゲラニル基(2水素付加)→テトラヒドロゲラニルゲラニル基(4水素付加)→フィチル基(6水素付加)となる。通常、植物が持つクロロフィルaを含め、多くの光合成生物のクロロフィル色素がフィチル基を持つ。合計3回の水素付加反応を単一の酵素であるゲラニルゲラニル還元酵素が担っている。
右:今回の研究によって、Halorhodospira halochlorisのゲラニルゲラニル還元酵素は、2箇所の還元反応しか行わない、4水素付加型の新規な反応特性を持つことがわかった。そのため、本菌のバクテリオクロロフィル最終産物はテトラヒドロゲラニルゲラニル基が結合したものである。
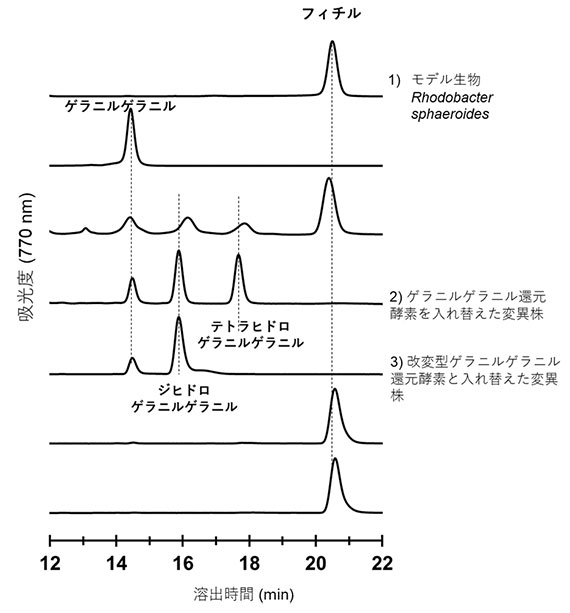
図3 高速液体クロマトグラフィーによる色素組成分析
4種類のイソプレノイド部位を持つバクテリオクロロフィル色素に由来する4つの溶出ピークが検出されている。溶出時間が早い順に、ゲラニルゲラニル基(14.5分)、ジヒドロゲラニルゲラニル基(16分)、テトラヒドロゲラニルゲラニル基(17.5分)、フィチル基(20.5分)を持つバクテリオクロロフィルに由来するピークである。
1) モデル光合成生物Rhodobacter sphaeroidesの色素組成。フィチル基がついたバクテリオクロロフィルのみを生産する。
2) ゲラニルゲラニル還元酵素を、Halorhodospira halochloris由来のゲラニルゲラニル還元酵素と入れ替えたRhodobacter sphaeroides変異株の色素組成。テトラヒドロゲラニルゲラニルまでしか還元が進んでいない。
3) ゲラニルゲラニル還元酵素を、アミノ酸の一部を改変したHalorhodospira halochloris由来ゲラニルゲラニル還元酵素と入れ替えたRhodobacter sphaeroides変異株の色素組成。ジヒドロゲラニルゲラニルまでしか還元が進んでいない。
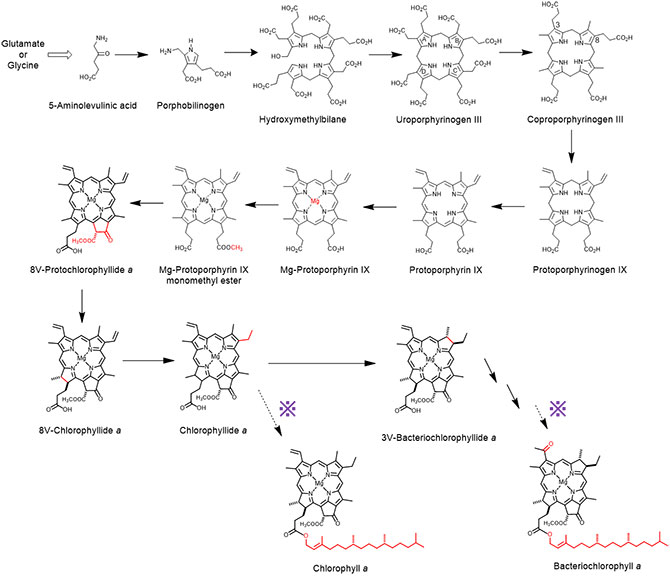
図4 クロロフィル色素類の生合成経路の模式図
クロロフィル色素は、アミノ酸派生物である5アミノレブリン酸を初発物質として、多段階の酵素反応ステップにより生合成される。生合成の最終段階(※印の破線矢印)において、クロロフィルかバクテリオクロロフィルかに関わらず共通のステップとして、ゲラニルゲラニル基の結合が起こり、次いでゲラニルゲラニル還元酵素によるゲラニルゲラニル基のフィチル化が起こる。
- (本研究について)
- 国立研究開発法人海洋研究開発機構
- 超先鋭研究開発部門 超先鋭研究プログラム
副主任研究員 塚谷祐介 - 久留米大学医学部医化学講座
- 講師 原田 二朗
- (報道担当)
- 国立研究開発法人海洋研究開発機構
- 海洋科学技術戦略部 報道室
- 立命館大学広報課