超深海(水深6000m以深の海溝環境、なお、太平洋には水深5,000~6,000mの深海平原が拡がります)の試料を採取するには、いくつかの方法があります。最も古典的な方法は、長いケーブルの先につけた採泥器や採水器を用いる方法です。とは言え、海洋学調査で最も普遍的に用いられる採水システム(CTDロゼット採水器)も、通常のケーブルでは、10,000mもの水深になるとケーブルが自重で切れてしまう可能性が高く、通常とは異なるチタンや特殊な繊維を使った特殊なケーブル(特注品)を準備する必要があるのです。その他、潜水艇を用いない方法として、自由落下で設置し、錘を外して浮上する江戸っ子1号のようなランダーシステムがあります。大がかりな設備は必要ないランダーシステムですが、実は浮上に必要なフロートが問題です。フロートも、10,000mの水圧に耐える必要があり、これも通常の深海調査で用いるフロートとは異なる特注品が必要です(工業スケールの特注品なので、そんな簡単に発注出来る代物ではありません)。
そして、最も一般に知られているのは有人あるいは無人の潜水船でしょう。超深海への最初の潜水船によるアクセスは、1960年のジャック・ピカールらによるマリアナ海溝の底へ「トリエステ」号(バチスカーフ)による探検です。この際、マリアナ海溝の底で、魚を見た!という、深海調査における伝説が生まれました。その後、有人潜水船による超深海への調査(探検)は、映画監督ジェイムス・キャメロンの一人乗り潜水船Deep Sea Challengerによる2012年の潜航調査まで、断絶しました。
一方、1995年、無人探査機による初の調査が、今は亡き初代無人探査機「かいこう」を用いて行われました。2003年に「かいこう」のビーグルが亡失した後に作られたのが、本研究に用いた大深度小型無人探査機「ABISMO」です。「ABISMO」は2007年に小笠原海溝、2008年にマリアナ海溝への潜航を成功させました。その後、2009年には、ケーブルありのROVモードと、自走式AUVモードを切り替えることが出来るウッズホール海洋研究所のHROV「ネレウス(Nereus)」がマリアナ海溝での調査に成功しています。2014年、「ABISMO」は引退が決定し、また、同年、「ネレウス(Nereus)」も調査中に失われました。つまり、現在、超深海へアクセス可能な現役の無人探査機は存在しない状況にあります。より詳細な歴史はHADES Programや、『The Hadal Zone: Life in the Deepest Oceans』(Alan Jamieson著)をご覧下さい。
超深海における微生物生態研究の歴史は1950年代に遡ります。当時は、船から降ろしたケーブルの先に付けた装置で採取した堆積物(つまり海底の泥)や、深海動物からの微生物の培養が試みられてきました。無人探査機「かいこう」によってマリアナ海溝から採取された試料からは単離培養の他、分子生態解析(環境試料から核酸を抽出して行う分子生物学解析)が試みられています。また、これまでの単離培養実験からは、高圧力下でのみ増殖可能な偏性好圧細菌の単離も報告されています。これらについては、上述の『The Hadal Zone: Life in the Deepest Oceans』(Alan Jamieson著)に纏められています。
そして、20世紀の超深海の微生物調査は、微生物生態研究というより、高圧下での生命活動の仕組みを知る、あるいは好圧細菌から有用な酵素等を探索することを目的として行われていました。超深海における本格的な微生物生態研究は、2013年のGludら(Nat Geosci誌)による、海溝底と周辺海域の堆積物中の微生物活性を比較すると、海溝底微生物活性が明らかに高いとする報告や、我々の小笠原海溝底堆積物中における無機窒素化合物循環と微生物活動の関係の報告まで待たねばなりません (Nunoura et al.2013. Environ.Microbiol誌)。
いずれにせよ、これまでの超深海の微生物調査においては、水塊試料は対象とされず、超深海・海溝水塊は未踏の海洋生命圏として残されていました。
今回の調査のきっかけは、初代「かいこう」亡失後も、水深10,000mの堆積物試料を確保したい(水深10,000mの堆積物を微生物単離のリソースとすることが、微生物部門の看板となっていた平和な時代の名残でした・・・)とするボスと、新技術を試すプラットフォームを作りたいとする開発部門の意向が合致したことに始まります。
とは言え、ただ堆積物を採ってきて、生態解析をやっても、普通の堆積物生態系やろ・・・という、醒めた目で見ていた我々は、どうせ新しいROVを作るなら、誰もやったことがない仕事が出来る設計にしましょ、と複数の深度で水塊試料を採取出来る設計を提案しました。そして、未踏の生態系であった海溝内水塊を、上層の水塊とも比較できるだけの採水能力を備えたROVの組み立てが始まったのです。つまり、今回の仕事の原型は、ここに誕生しました。
2007年の小笠原海溝調査では採泥のみでしたが、2008年にはマリアナ海溝チャレンジャー海淵等で採水・採泥に成功しました。その後、2011年に小笠原海溝及びその周辺にて、採水・採泥を行いましたが、この航海が「ABISMO」による最後の10,000m級潜航でした。2014年に再度マリアナ海溝にて水深7,900mまでの潜航を行い(ROVケーブルが老朽化し、これ以上潜れなくなっていました・・・、ケーブルは特注品であり、そんな簡単には作れません)、これが引退興行となりました。
従来、海洋表層で珪藻やシアノバクテリア等が光合成により生産された有機物が、日光の到達しない深海環境では、微生物や動物プランクトンに分解されつつ沈降し、最終的には深海域の生命圏を支えるというモデルが一般的に信じられてきました。つまり、深海水塊に棲息する微生物の殆どは従属栄養生物であると認識されており、深海で日光に依存せずに炭素固定を行う化学合成生物は、熱水環境等、限られた環境にのみ生息すると考えられていたのです。
ところが、近年の研究は、海洋表層で生産された有機物の分解等に伴って生じるアンモニア(有機窒素化合物の分解から生じる)や硫黄化合物(マリンスノー等の中に生じる微小な嫌気環境での硫酸還元から生じる硫化水素や、(微)生物に由来する有機硫黄化合物等)を酸化してエネルギーを得て炭素固定する微生物が、深海水塊中にも普遍的に分布することが明らかになりました。それだけでなく、アンモニア酸化アーキアなどは、深海水塊中の微生物生態系において相当程度優占することも示唆されています。
即ち、最新の深海微生物生態系モデルでは、海洋表層で生産された有機物はただ分解されて沈降するのではなく(図中では沈降有機物:デトライタスと表現)、有機物の分解と合成を繰り返しながら海底に至るという新しい生態系像が構築されつつあります。ただし、水塊中で暗黒の炭素固定を行う微生物が、炭素固定に完全に依存しているのか、あるいは、有機物摂取と炭素固定を共存させる混合栄養戦略をとっているのか、まだ明らかになっていません。詳細は、『生物の科学 遺伝』2014年11月号に紹介されています。
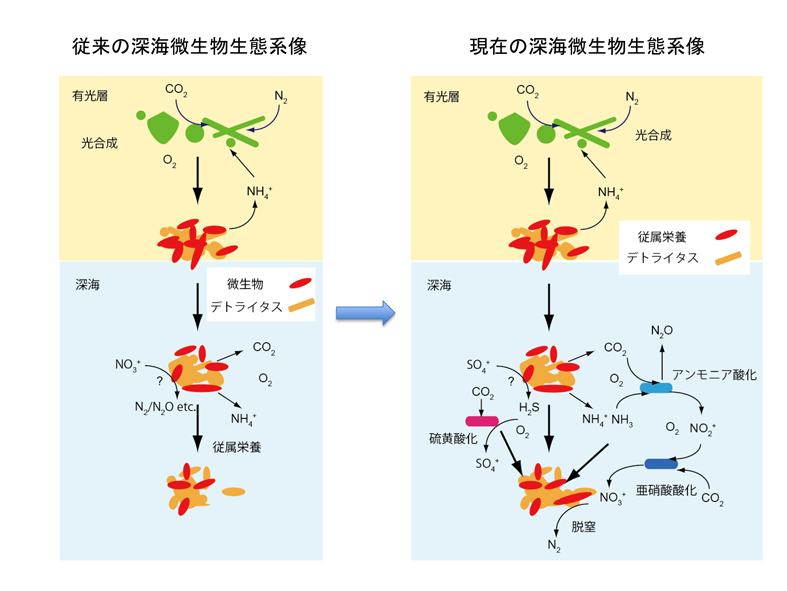
原核生物(バクテリア・アーキア)のSSU rRNA遺伝子を対象とした多様性解析では、超深海と深海層では、微生物群集構造に明瞭な差異が確認されました。そして、その群集構造の違いは、超深海には従属栄養系統群(有機物を炭素・エネルギー源とする)が優占し、深海層では炭素固定能を有す系統群が優占することを示しています。
その一方、化学・物理データは、超深海と深海層での明瞭な違いを示していません。特に、微生物による有機窒素化合物の分解で生じるアンモニアについては、非常に濃度が低いため、正確な濃度測定すら難しく、今回の調査では、確かなデータを得ることすらできませんでした。なお、この場合、アンモニアの濃度が非常に低い原因として、微生物による生産と消費が拮抗しているため、生産量(あるいは移入量)と消費量の差である現場での蓄積(すなわち濃度)は、検出できなかったと考えられます。
そこで、アンモニアをエネルギー源とするアンモニア酸化菌(亜硝酸菌とも書かれます)、アンモニア酸化菌の生産する亜硝酸をエネルギー源とする亜硝酸酸化菌(硝酸菌とも書かれます)の棲み分けを利用して現場における有機物分解の傾向を検証することにしました。
アンモニア酸化菌では単一系統群のアンモニア酸化アーキア(AOA)と複数系統群のアンモニア酸化バクテリア(AOB)が知られており、AOBの方がAOAに比べ、高濃度のアンモニアを好むこと、また、AOAの中でも複数のグループが、それぞれ異なった至適アンモニア濃度(フラックス)を有することが、これまでの研究によって示唆されています。亜硝酸酸化菌には複数のバクテリア系統群があり、同様に至適亜硝酸濃度(フラックス)が異なる系統群が知られています。
本研究では、超深海水塊において、それぞれ比較的高濃度のアンモニア、亜硝酸を好む系統群が優占すること、それに対し、深海層では低濃度の基質を好む系統群が優占することが明らかになり、微生物群集全体の変化からの解釈と合致する結果を得ることができました。なお、海洋表層においては、基質濃度だけでなく、光に対する感受性も棲み分けに影響を及ぼしていると考えられています。背景となる詳細な知見については、『生物の科学 遺伝』2014年11月号や本論文に挙げた参考文献をご覧ください。
注)アンモニアから硝酸に至る過程をまとめて硝化、また、アンモニア酸化菌と亜硝酸酸化菌を合わせて硝化菌とも呼びます。特にAOAの棲み分けについては、現場環境におけるアンモニア、亜硝酸濃度の測定がしばしば検出限界より低いこと、また、低濃度を好む系統群については、培養による評価が困難であることから、硝化菌群が利用可能な供給量という意を含めフラックスという言葉で表現しています。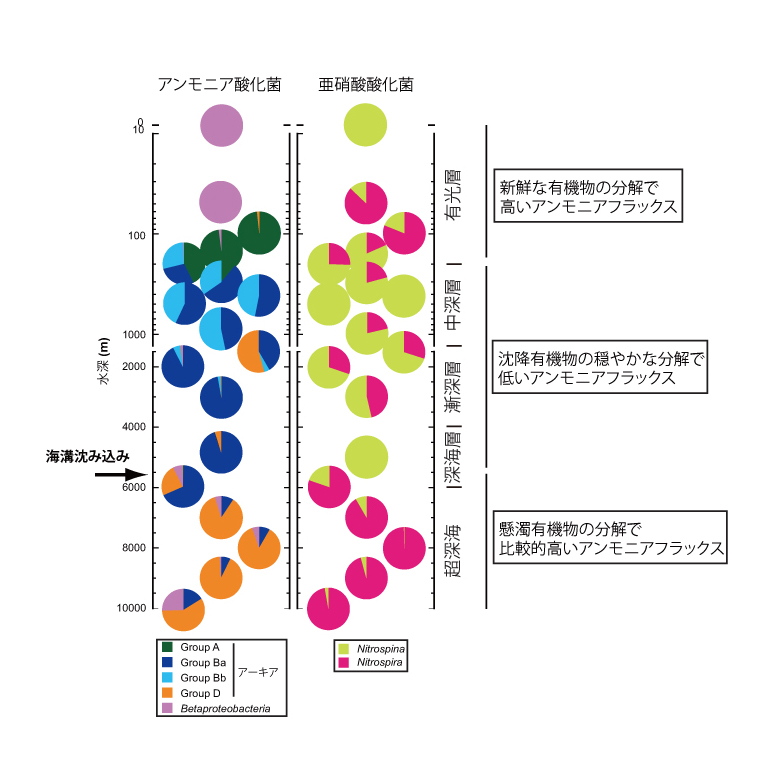
深い海には原始的な生命が存在するだとか、深いところには特別な微生物が存在する、という説明を聞いた記憶のある方は多いと思います。今回の得られた結果から言えることは、少なくともバクテリア・アーキアに関しては、原始的な系統群、特別な系統群は存在せず、普通の海洋微生物群が棲んでいます。我々の先人は、過剰な表現をしてきました、代わって謝罪します、ゴメンナサイ、ということです。少なくとも、未知の原始的な系統群は検出できませんでしたし、検出された微生物系統群が示す微生物生態系の構造も、普通の海洋環境のそれと本質的に異なるようなものではありませんでした。
ただし、ここでの系統群とは微生物分類での属レベルより高次の系統群レベルでの話です。これまでの好圧細菌の単離が示すように、種あるいは株レベルにおいて圧力環境に適応したものは存在します。つまり、本質的に高圧環境下で適応してきた系統群が存在するのではなく、個々の微生物系統において比較的容易に高圧環境に適応していると考えられます。また、有孔虫では海溝底からは始原的な系統が見つかっています(Todo et al. 2005, Science誌)ので、真核生物の個々の分類群レベルでは、超深海に原始的な種が生息する傾向はあるのかもしれません。
そもそも、圧力の生物組織へ与える影響とは何でしょうか? インスタントヌードルの発泡スチロール容器が高水圧下で収縮するのは、発泡スチロールの空隙中の空気が収縮すると共に、空隙を支える構造が圧力により崩れる為です。逆に、空気の入ったゴム風船のように、気圧が低くなると、さらに膨らんで破裂する、ということもあります。生物についても同様であり、圧力変動時の構造の維持が課題になります。我々のような動物の体は、分子(タンパク質・脂質等)― 細胞 ― 組織(臓器等)― 動物(生体)という階層で出来ています。圧力は個々の段階での脆弱性に効きますから、より複雑な構造を有する生物ほど、圧力の異なる環境への適応は難しいと言えるでしょう。
今回の研究の対象となる単細胞のバクテリア・アーキアであれば、生体を構成する分子そのものが望ましい構造を維持できるかどうかが最大の焦点となります。また、生物は通常、急激に圧力の異なる環境へ移動することはありません。つまり、個々の生息環境における遺伝子の中立進化(遺伝子レベルの微細な変異の多くは、生存に有利にも不利にも働かないと考えられています)により、偶々、生存に不可欠な分子が、高圧力下で分子の構造を保てなくなったものが圧力感受性菌となり、また、低圧力下で分子構造を保てなくなったものが好圧菌となる、と考えるのが最もリーズナブルです。
布浦拓郎(海洋生命理工学研究開発センター)
