プレスリリース
2016年 10月 11日
国立研究開発法人海洋研究開発機構
国立大学法人東京大学大学院理学系研究科
国立大学法人九州大学
国立大学法人高知大学
沖縄熱水海底下生命圏掘削により熱水孔下生命圏の限界を発見
~海底下の変動する高温環境が生命圏の限界を決定する~
1.概要
国立研究開発法人海洋研究開発機構(以下「JAMSTEC」という。)深海・地殻内生物圏研究分野の柳川勝紀ポストドクトラル研究員(当時、現九州大学大学院比較社会文化研究院学術研究員)らは東京大学、九州大学、高知大学、ドイツ連邦地球科学天然資源研究所と共同で、2010年9月に実施した中部沖縄トラフ熱水活動域での科学掘削(沖縄熱水海底下生命圏掘削:2010年10月5日既報)で取得した柱状試料(コアサンプル)を用いて、深海熱水噴出孔近傍の海底下に存在する生命圏「熱水噴出孔直下生命圏」の存在様式とその限界を明らかにしました。
これまでの深海熱水活動域の噴出熱水や熱水チムニー等を対象とした研究により、熱水噴出孔の海底下には地球内部エネルギー(還元的無機化学物質)に依存した超好熱性微生物(※1)の住処(熱水噴出孔直下生命圏)があることが強く予見されていました。しかし、科学掘削やその試料による直接的な証明に到った研究例はありませんでした。統合国際深海掘削計画(IODP※2)第331次研究航海では、この熱水噴出孔直下生命圏の直接的証明とその実態を世界に先駆けて解明することを目的に、中部沖縄トラフ伊平屋北海丘において地球深部探査船「ちきゅう」による掘削調査を行いました。本研究では、その航海で取得されたコアサンプルの詳細な地球化学的分析と微生物学的解析を行い、微生物学、地球化学、鉱物学的手法を組み合わせた学際統合的な手法と解釈を通じて、海底下高温熱水溜まりの変動やその化学組成の変化に伴う生息環境の空間的・時間的変化に規制された「熱水噴出孔直下生命圏」の存在を明らかにすることができました。また、本研究は、海底下環境に生命圏と非生命圏の境界が存在することを世界で初めて明確に捉えることに成功し、その生息限界を決定づける物理・化学条件(今回の場合は高温)を明らかにした点で、地球における生命活動や生命圏の限界という根源的な科学命題に一つの答えを与える画期的な成果と言えます。
IODP第331次研究航海は、これまで掘削試料や掘削後調査を通じて、海底熱水鉱床研究開発やその成因に対する重要な科学的知見を提示するだけでなく(2010年10月5日既報および2016年2月25日既報)、科学掘削による環境変化が熱水化学合成生物群集に及ぼす影響の世界初の定量的評価(2015年4月23日既報)や今後の海底工学やエネルギー資源研究における新しい可能性(2013年9月3日既報)を提示するなど、多くの研究成果を挙げてきています。これら成果の原点である、本研究成果こそがIODP第331次研究航海の科学目標を達成すべき最も重要な成果と言えます。
なお、本成果は、国際微生物生態学会が発行する科学誌のThe ISME Journal(Nature Publishing Group)のオンライン版に10月11日付け(日本時間)で掲載される予定です。
タイトル:Defining boundaries for the distribution of microbial communities beneath the sediment-buried, hydrothermally active seafloor.
著者:柳川勝紀1,2,3、井尻暁4、Anja Breuker5、酒井早苗1、三好陽子6、川口慎介1、野口拓郎7、平井美穂8、Axel Schippers5、石橋純一郎6、高木善弘1、砂村倫成2、浦辺徹郎2、布浦拓郎8、高井研1
所属:1.海洋研究開発機構 深海・地殻内生物圏研究分野、2.東京大学大学院理学系研究科、3.九州大学大学院比較社会文化研究院、4.海洋研究開発機構 高知コア研究所地球深部生命研究グループ、5.ドイツ連邦地球科学天然資源研究所、6.九州大学大学院理学府、7.高知大学教育研究部総合科学系複合領域科学部門、8.海洋研究開発機構 海洋生命理工学研究開発センター
2.背景
深海熱水活動域には、周辺海洋環境とは異なる特異的な微生物群集が見られます。特に、熱水由来の還元的な無機化学物質を利用できる化学合成独立栄養微生物(※3)や高温環境での生育を好む超好熱性微生物など、熱水域の物理・化学条件に強く依存した微生物からなる生態系が存在しています。これらの微生物生態系は、熱水噴出孔直下の海底下に広がるそのような微生物群の「巣」(熱水噴出孔直下生命圏)から、一部の「タネ」が熱水によって海底にもたらされ群集を形成した結果であるという有力な仮説が知られていました。これまでも「熱水噴出孔直下生命圏」を直接的に証明するために、深海熱水活動域の掘削が国内外で複数回実施されてきましたが、掘削試料の回収率が低いことや周辺環境からの微生物汚染等の技術的問題から、熱水噴出孔直下生命圏の証明ならびに実態解明には到りませんでした。
2010年9月に、深海熱水活動域直下における微生物群集の存在様式および生態系を世界に先駆けて解明することを目的として、中部沖縄トラフ伊平屋北熱水活動域(図1)において「ちきゅう」による科学掘削を実施しました(2010年9月1日および同10月5日既報:統合国際深海掘削計画(IODP)第331次研究航海「沖縄熱水海底下生命圏掘削-1」)。この航海では計5地点で掘削が行われていますが、掘削後に形成された新しい熱水噴出孔の出現パターンや掘削で測定された温度構造(図2)を考慮した場合、熱水活動中心から450mほど離れた掘削地点C0014が、熱水噴出孔直下生命圏研究に最適な環境であると考えられました。そこで、本研究では掘削地点C0014での掘削試料を対象とした微生物学、地球化学、鉱物学的手法を組み合わせた分野横断的研究を実施しました。
3.成果
沖縄トラフの深海熱水活動域で噴出する高温熱水にはカリウムなどのアルカリ元素や二酸化炭素、メタンといったガス成分が多量に含まれるという化学的特徴が知られています。取得したコアサンプルの間隙水にもその熱水の化学的特徴は観察され、例えば掘削地点C0014の1つの掘削孔(Hole B)では、ある深度からカリウム濃度の急激な上昇が測定されています(図3)。一方で、メタンでは3つの濃度異常が確認され、気相に富んだ熱水(あるいはその影響)が局所的に存在していることが分かりました。このことは海底下で化学組成の異なる複数の熱水流や熱水滞留が存在し、浸透する海水と混ざり合いながら、複雑な水理構造が発達していることを示唆しています。
海底下8.5 mではメタンの炭素同位体比が顕著に大きいことから、微生物による活発なメタン消費が予想されました(図3)。このことは、放射性同位体トレーサーを用いた活性測定実験(※4)によって明らかとなった活発な嫌気的メタン酸化活性(※5)とも調和的でした(図4)。さらに、嫌気的メタン酸化反応の鍵となる酵素の遺伝子(mcrA遺伝子※6)の組成と量を分析したところ、高温環境を好む嫌気的メタン酸化アーキア(古細菌※7)由来のmcrA遺伝子が多く検出されました(図4)。これらのことは、高温適応型のアーキアによるメタンの消費が、伊平屋北海丘の熱水噴出孔直下生命圏の特徴の1つであることを示します。
培養による検出、蛍光顕微鏡下での観察、定量PCR法(※8)による16SリボソームRNA遺伝子の定量、放射性同位体トレーサーを用いた有機物分解活性といった複数の方法を用いて、綿密に微生物の検出を行った結果、興味深いことに、この掘削孔において海底下14.3m以深では全く微生物が存在していない環境が広がっていることがわかりました(図5)。また、16SリボソームRNA遺伝子配列に基づく微生物群集組成では、表層からは一般的な海洋堆積物に存在するバクテリアが多数検出されました。一方定量PCR法の結果は、微生物が存在しうる領域(生命圏)の最深部ではアーキアが相対的に多く、微生物群集の半数を占めることを示していました(図5)。実際に、最深部の試料からはHot Water Crenarchaeotic Group IV(HWCGIV)という未培養系統群に属す好熱性アーキア由来の16SリボソームRNA遺伝子が検出されました。このアーキアは他の深海熱水環境からも報告されており、熱水の化学成分を利用した微生物であると考えられ、16SリボソームRNA遺伝子の塩基配列を利用した生育温度推定法によってその至適生育温度は50-76 °Cと予想されます。一方、80 °Cを超える環境で生育するような超好熱菌は、培養法でも16SリボソームRNA遺伝子を対象とした分子生物学的手法でも検出されませんでした。熱水噴出孔直下生命圏では生命の最高生育温度(122°C、2008年 7月 29日既報)に近い温度で生育可能な微生物が存在することが予見されていましたが、今回の結果はこれに反するものでした。その理由として、過去の海底下高温熱水の海底下水位の増減(つまり海底下環境の温度履歴)が考えられ、堆積層内に見られる熱水変質鉱物(※9)に着目したところ、現在の生命圏と非生命圏の境界に当たる深度において高温条件下で生成されたと考えられる熱水変質鉱物を確認することができました。さらに、これらの熱水変質鉱物の酸素同位体比に基づく温度推定から、この境界が過去に106°C以上の高温環境に曝されていたことが示されています(図6)。このことは、かつて存在していた生命圏の深部に高温熱水が流入、あるいは高温熱水溜まりの水位が上昇したことで、生息していた微生物が全滅し、その後に熱水が流出あるいは水位が低下したことで温度条件が生存可能となり、現在のHWCGIVが優占する微生物生態系(生命圏)が回復しつつある状況であると考えられます。
4.今後の展望
今回の沖縄熱水海底下生命圏掘削研究において,世界で初めて海底下環境に生命圏と非生命圏の境界を発見し、その生息限界を決定する温度条件を明らかにしたことによって、地球における生命活動や生命圏の限界という根源的な科学命題に一つの答えを与えることができました。さらに、本研究では、熱水噴出孔直下生命圏の存在やその規模・構造は過去の熱水活動履歴に強く依存しているということが示され、図らずもダイナミックに変動する熱水活動とそれに依存した微生物生態系の関係性を知ることができました。地球上での生命誕生の場所と考えられる熱水活動域における生命圏の存在形の一つを明らかにできたことは、今後の生命誕生の場を考える上で、非常に重要な知見となります。一方、「ちきゅう」研究航海前に予想していた「多様な超好熱性アーキアの巣」や「熱水孔直下に潜む原始的生命」を発見することはできませんでしたが、それらの存在は否定されたわけではありません。そのためには、沖縄トラフの熱水活動域とは異なる型の熱水活動域、つまり海底下熱水の移流や滞留層が比較的安定した海底熱水環境、例えば中央インド洋海嶺のかいれいフィールドのような堆積物に覆われていない水素に富んだ中央海嶺型熱水域を調べることが必要です。海底下熱水の移流や滞留層の変動によって生命圏が規定される過程では、熱水の挙動や微生物の移動を支配する不透水層(※10)が重要な役割を果たします。この不透水層は海底熱水鉱床の成長にも大きな影響を与えることがわかりつつあります。不透水層の広がりは、海底下熱水鉱床の成因(特にその規模の拡大)に大きな影響を与えます。よって不透水層とその下に広がる海底下熱水溜まりの存在を可視化し、評価することは、海底下生命圏の広がりだけでなく、海底下熱水鉱床の分布を見積もることにも結びつきます。すなわち海底下熱水鉱床の研究開発を通じて海底下生命圏の理解が進むことも期待できます。
※1 超好熱性
80°C以上の高温に至適生育温度を有する生理学的特徴。
※2 国際深海科学掘削計画(IODP: International Ocean Discovery Program)
平成25年10月から開始された多国間科学研究協力プロジェクト。日本(地球深部探査船「ちきゅう」)、アメリカ(ジョイデス・レゾリューション号)、ヨーロッパ(特定任務掘削船)がそれぞれ提供する掘削船を用いて深海底を掘削することにより、地球環境変動、地球内部構造、海底下生命圏等の解明を目的とした研究を行う。なお、本プロジェクトは平成15年10月から平成25年まで実施された統合国際深海掘削計画(IODP:Integrated Ocean Drilling Program)から引き継いでいる。
※3 化学合成独立栄養微生物
水素、硫化水素など還元的無機物質を酸化することでエネルギーを得るとともに、そのエネルギーによって二酸化炭素を有機物に変換することで生育する微生物。
※4 放射性同位体トレーサーを用いた活性測定実験
任意の微生物代謝で使われる基質に放射性同位体標識された化合物を加え、反応産物の放射線強度を測定することで、単位時間あたりの代謝活性を求めることができる。本研究では14C標識二酸化炭素(14CO2)やメタン(14CH4)等を掘削試料に添加し、一定期間培養後のメタン(14CO2→14CH4)や二酸化炭素(14CH4→14CO2)等の微生物活性を計測とした。
※5 嫌気的メタン酸化
メタンを嫌気的条件下で二酸化炭素に変換する反応。この反応は嫌気的メタン酸化アーキア(ANME)が担うと考えられており、硫酸還元菌と共生することが一般的に知られる(CH4 + SO42-→HCO3-+HS-+H2O)。
※6 mcrA遺伝子
メタン生成アーキアでは、メタン生成の最終酵素反応、嫌気的メタン酸化アーキアでは、メタンを資化する最初の酵素反応をになうMethyl CoM reductaseの大サブユニットをコードする遺伝子
※7 アーキア(古細菌)
地球上のあらゆる生命体は、アーキア(古細菌)・バクテリア(真正細菌)・ユーカリア(真核生物)の三つに分類される。微生物の多くはアーキアとバクテリアに属しているが、両者は、細胞膜を構成する極性脂質の構造や遺伝子の転写システム等、生命の根幹に異なる生化学的、分子生物学的性質が異なるため区別することができる。
※8 定量PCR法
定量PCR(Quantitative polymerase chain reaction)。特定の遺伝子をポリメラーゼ連鎖反応で増幅し、その増幅産物の増幅速度を定量することで、増幅前の試料中の遺伝子数を算出する方法。試料中の特定の代謝機能を有する微生物群の存在量や活動度を推定することができる。本研究では、16SリボソームRNA遺伝子やmcrA遺伝子などの遺伝子数を定量し、微生物全体やメタン生成アーキア、嫌気的メタン酸化アーキアの存在量を推定した。
※9 熱水変質鉱物
岩石を構成する鉱物が熱水との反応により、異なる鉱物に変化したもの。
※10 不透水層
地層を構成する粒子の間隙が狭く、水を通さない(通しにくい)地層。熱水変質作用で粘土鉱物が大量に生成されると不透水層になりやすい。
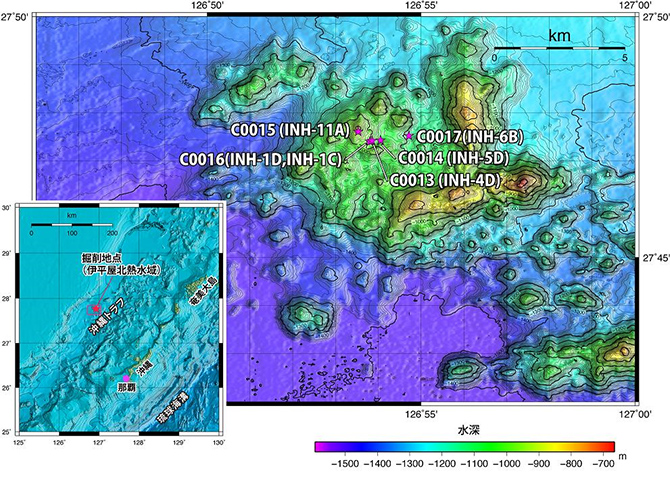
図1 調査海域
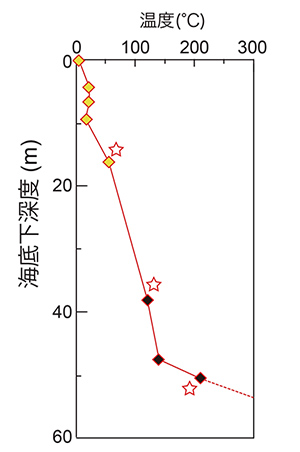
図2 掘削地点C0014における温度勾配(Takai et al., 2011を改変)。黄色のプロットはコアリングと並行して温度計測装置(APCT-3)で測定されたもので、黒色はコアライナーチューブに貼られた示温テープから見積もった値を示す。星印のあるものは下限値を示している。
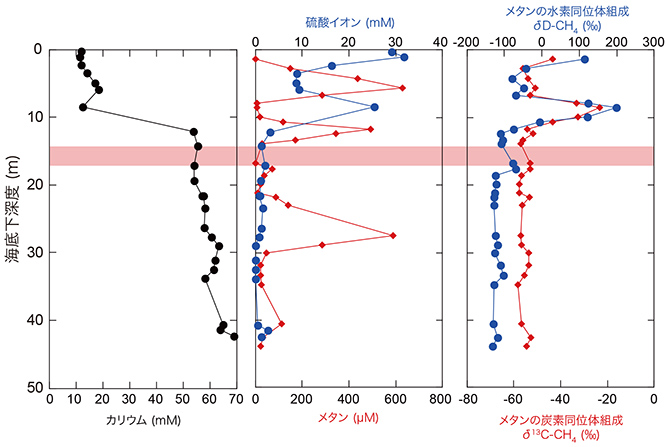
図3 掘削地点C0014 Hole Bでの間隙水化学組成のプロファイル。カリウムは深さ10 mから急激に増加するが、メタンは3つのピーク(5、11、27m)を持つ。メタンの炭素・水素同位体組成は共に深さ8.5mで顕著に高くなるが、これは微生物によるメタン消費が盛んであることを示唆している。なお、生命圏限界に相当する深度を赤色帯で示した。
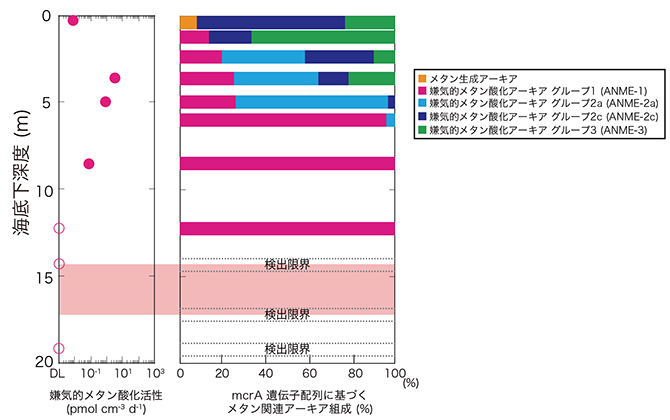
図4 掘削地点C0014 Hole Bでの嫌気的メタン酸化活性とメタン生成/嫌気的メタン酸化アーキアの群集組成。嫌気的メタン酸化活性でy軸上の白抜きのプロットは検出限界値以下を示す。メタン生成/嫌気的メタン酸化アーキアの群集組成は、各反応の鍵酵素をコードする遺伝子(mcrA)の塩基配列をもとに決定された。生命圏限界に相当する深度を赤色帯で示した。

図5 掘削地点C0014 Hole Bでの生命圏分布と微生物群集組成。左から、(1)全菌数と16SリボソームRNA遺伝子数、(2)酢酸分解活性、(3)原核生物由来16SリボソームRNA遺伝子数とアーキア由来16SリボソームRNA遺伝子数の比、(4) 16SリボソームRNA遺伝子に基づく微生物群集組成。y軸上の白抜きのプロットは検出限界値以下を示す。微生物群集組成で星印の深度では、アーキア特異的プライマーを用いた場合にのみ遺伝子増幅が可能であった。生命圏限界に相当する深度には赤色帯で示した。
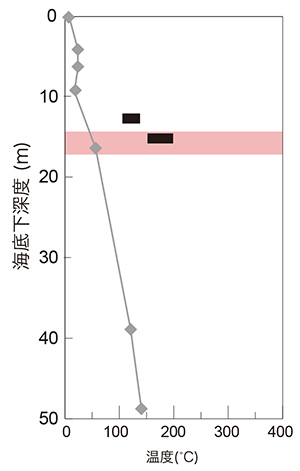
図6 掘削地点C0014の海底下温度の測定値と推測値の比較。灰色で図2に示した温度プロファイルを示した。黒い太線は、堆積層内の熱水変質鉱物の生成温度について、その同位体比組成から推定された温度範囲を示す。生命圏限界に相当する深度は赤色で示した。
沖縄熱水域の海底下に生命圏の限界を見た!
- (本研究について)
- 国立研究開発法人海洋研究開発機構 深海・地殻内生物圏研究分野
- 分野長 高井 研
- 国立大学法人東京大学 大学院理学系研究科
- 助教 砂村 倫成
- 国立大学法人九州大学
- 大学院理学研究院 准教授 石橋 純一郎
- 大学院比較社会文化研究院 学術研究員 柳川 勝紀
- 国立大学法人高知大学 教育研究部総合科学系複合領域科学部門
- 准教授 野口 拓郎
- (報道担当)
- 国立研究開発法人海洋研究開発機構
- 広報部 報道課長 野口 剛
- 国立大学法人東京大学大学院理学系研究科
- 広報室 特任専門職員 武田 加奈子、教授・広報室長 山内 薫
- 国立大学法人九州大学
- 広報室
- 国立大学法人高知大学
- 総務課広報係 石田