プレスリリース
国立研究開発法人海洋研究開発機構
国立大学法人高知大学
国立研究開発法人産業技術総合研究所
国立大学法人東京大学
国立大学法人京都大学
国立大学法人神戸大学
深海掘削により室戸岬沖の海底下生命圏の実態とその温度限界を解明
1. 発表のポイント
- ◆
- 南海トラフ沈み込み帯先端部の海底堆積物環境において、40-50℃と70℃付近の深度区間が、生命(微生物)の存続にとって重要な温度限界域であることを突き止めた。
- ◆
- 海底下生命圏の温度限界域に、微生物の生存戦略の一つの形態である内生胞子が高濃度に存在することを見出した。
- ◆
- 70℃付近と90-110℃の深度区間に、微生物細胞や代謝活動のシグナルが検出されない環境を認めた。その深度区間には、微生物の消費を免れた高濃度の酢酸が存在していた。
- ◆
- 110-120℃の堆積物―基盤岩境界域に、酢酸を消費する超好熱性微生物生態系の存在を発見した。
2. 概要
国立研究開発法人海洋研究開発機構(理事長 松永 是)研究プラットフォーム運用開発部門 マントル掘削プロモーション室の稲垣史生室長と超先鋭研究開発部門 高知コア研究所 地球微生物学研究グループの諸野祐樹主任研究員らは、ブレーメン大学やロードアイランド大学、高知大学などと共同で、地球深部探査船「ちきゅう」を用いて高知県室戸岬沖の南海トラフ沈み込み帯先端部の海底(水深4776 m・1.7℃)から深度1180 m・120℃までの堆積物コア試料(※1)を採取し、海底下環境に生息する微生物の分布や間隙水中の化学成分、堆積物の物性や温度などを詳細に分析しました。その結果、室戸岬沖の地質環境と温度条件に依存した海底下生命圏の実態とその限界が明らかになりました。
これまでの研究により、地質学的時間スケールをかけて形成される海底堆積物には、未だ培養されていない固有の微生物が生息していることが明らかとなっています。それらの微生物活動は、海水から埋没した有機物の分解やメタンハイドレートの形成など、地球規模の物質循環に重要な役割を果たしていると考えられています。しかし、海底下のどの程度の深さまで生命圏が拡がっているのか、その限界を規定する環境要因とは何か、といった根本的な疑問が未解明のまま残されていました。
それらの科学的疑問を解き明かすことを目的として、2016年に国際深海科学掘削計画(IODP、※2)第370次研究航海「室戸沖限界生命圏掘削調査:T-リミット」が、地球深部探査船「ちきゅう」と高知コアセンター(※3)の研究施設を用いて実施されました。その結果、堆積物上部の低温〜常温環境に生息する微生物細胞の密度は、温度(深度)が増すにつれて低下し、約45℃以上では1 cm3あたり100細胞以下にまで減少することが明らかとなりました。また、40-45℃の深度区間と75-90℃の深度区間では、内生胞子(※4)の密度が局所的に増加している現象が認められました。一方、45℃以上の高温環境では、プレート境界断層(※5)下部の90-100℃の環境に、微生物細胞が検出されない深度区間が存在し、微生物による消費を受けないため高濃度の酢酸が存在することが明らかとなりました。また、それより深い110-120℃の堆積物―基盤岩境界域には、酢酸からのメタン生成を示す化学成分濃度や炭素同位体組成の変化や、細胞密度の増加など、超好熱性微生物生態系の存在を示す複数の特徴が認められました。
本研究で得られた知見は、現場の温度や栄養・エネルギー状態のみならず、室戸岬沖の南海トラフ沈み込み帯先端部における地質学的プロセスや流体移動プロセスが、海底下深部環境における生命生息可能条件(ハビタビリティ)に重要な影響を与えていることを示しています。また、海洋プレートの沈み込み帯において、120℃までの堆積物―基盤岩境界域においてもなお生命シグナルが検出されたことから、地球惑星における生命圏の広がりとその限界の可能性は、海洋プレートが沈み込むその先や堆積物の下に広がる岩石圏(海洋地殻や上部マントル)にまで及ぶことが示唆されました。
本研究の一部は、日本学術振興会の最先端研究基盤事業、最先端・次世代研究開発支援プログラム(GR102)、科学研究費助成事業(JP26251041、JP19H05503、JP20K20429、JP19H00730)、ドイツ研究振興協会(DFG)、アメリカ国立科学財団(NSF)、英国自然環境研究会議(NERC)、アルフレッド・P・スローン財団(Deep Carbon Observatory)の支援を受けて行われたものです。
本成果は、アメリカ科学振興協会(AAAS)が発行する科学誌「サイエンス」のオンライン版に12月4日付け(日本時間)に掲載される予定です。
- ブレーメン大学(ドイツ)
- 国立研究開発法人海洋研究開発機構
- ロードアイランド大学(米国)
- カリフォルニア州立大学ロサンゼルス校(米国)
- オーフス大学(デンマーク)
- 国立大学法人高知大学
- アバディーン大学(英国)
- カルガリー大学(カナダ)
- アルフレッド・ウェゲナー極地研究所(ドイツ)
- 学校法人日本大学(現・京都大学)
- 国立研究開発法人産業技術総合研究所
- スイス連邦工科大学(スイス)
- プリマス大学(英国)
- ワシントン大学セントルイス(米国)
- カリフォルニア工科大学(米国)
- 国立大学法人東京大学
- クイーンズランド大学(オーストラリア)
- トロント大学(カナダ)
- マサチューセッツ工科大学(米国)
- テキサスA&M大学(米国)
- 国立研究開発法人海洋研究開発機構(現・神戸大学)
- ヨンセイ大学(韓国)
- GFZヘルムホルツセンターポツダム(ドイツ)
(*これらの著者は、この研究に同等に貢献しています。†責任著者。)
3. 背景
これまでの海洋科学掘削調査により、地球表層の約7割を占める広大な海洋のその下に、膨大な数の生命(微生物)が存在することが明らかとなっています。その微生物細胞の数は、地球全体で2.9-5.4×1029細胞であると推定されています。海底下には、推定4万種におよぶ性状未知の微生物が存在し、陸域の土壌や海水中に生息する微生物と同等の生物多様性があることが明らかとなっています(2020年10月20日既報)。それらの海底下微生物を構成する炭素は、地球に存在する全ての生物の炭素の約0.18-3.6%にしか過ぎません。しかし、近年の研究により、海底下の微生物は、海底下の限られた栄養・エネルギー条件下であっても、1億年以上もの地質学的な時間を生き抜くことが実証されています(2020年7月29日既報)。それらの海底下微生物の現場における代謝活性は、一つの細胞が1日あたりに数十個の電子しか使わない程度の微弱なものです。そのため、海底下の過酷な低エネルギー環境に適応した微生物には、生命機能を維持するのに最低限のエネルギーを獲得しつつ、地質学的な時間スケールでの存続を可能とする未知の生理・代謝機能があると考えられています(2011年10月11日既報)。
一般的に、海底堆積物やその下の海洋地殻環境は、時間が経過し、深くなるにつれて温度が上昇することが知られています。また、地球上の海底堆積物の体積(3.0×108 km3)において、40℃以下の堆積物が占める割合は約48%(1.4×108 km3)であり、40℃以上の温度の高い堆積物とおよそ同じ割合であると考えられています。時間とともに変化する地層の温度に対して、海底下の微生物はどのように適応・分布しているのでしょうか?2012年、私たちは地球深部探査船「ちきゅう」を用いて下北半島八戸沖の海底を2466 m掘削し、推定温度63.7℃までの全ての堆積物環境に、有機物を分解しメタンを生成する従属栄養型の微生物生態系が存在していることを突き止めました(2015年7月24日既報)。下北沖の海底下では、深度約1.2-1.6 km(温度40-45℃)付近から微生物細胞密度の急激な低下が認められたことから、海底堆積物に生息する微生物の存在量や分布は温度と関係し、その代謝活性は、地質環境が定める現場の栄養・エネルギー供給に依存しているのではないかという仮説が立てられました。
それらの仮説を実証するため、2016年9月、高知県室戸岬の沖合約125 kmの地点(サイトC0023:水深4776 m、図1)において、南海トラフ沈み込み帯先端部における海底下生命圏の温度限界の実態解明に向けた国際深海科学掘削計画(IODP)第370次研究航海(通称「T-リミット」)が実施されました(2016年9月5日既報)。本研究航海の調査海域は、過去に海洋科学掘削による地質学的な調査が行われており、比較的地温勾配が高く、深度約1.2 kmの基盤岩の温度は超好熱性微生物の温度限界に相当する120℃付近に達することが予想されました。本IODP研究航海では、「ちきゅう」により掘削採取されたコア試料を船上のX線CTスキャンで分析し、微生物分析用のコア試料を船上で分取した後、それらのサンプルをヘリコプターで高知コアセンター(高知県南国市)に輸送し、クリーンルーム内で無菌的な(外部汚染のない)処理を施しました。また、掘削された孔内の複数箇所に温度計を設置し、2018年3月に行われた海洋研究開発機構による深海調査研究船「かいれい」と無人探査機「かいこう」を用いた深海調査(KR18-04)において観測温度データの回収を行いました。
4. 成果
「ちきゅう」の掘削により直接的に測定された温度データと堆積物コア試料の熱伝導率の測定データに基づき、掘削孔最深部(深度1180 m)の温度は120±3℃であることが示されました。また、2018年に実施された深海調査により、「ちきゅう」により設置された孔内温度観測のデータが回収され、本温度データの有効性が確認されました。
海底下約190-400 m・30-50℃までの深度区間において、深度・温度が増加するにつれて微生物細胞の密度が低下し、蛍光染色法による微生物細胞密度の定量限界値(1 cm3あたり16細胞)以下にまで減少する傾向が認められました(図2A)。対照的に、バクテリアの内生胞子の密度が増加し、深度400 m付近で1 cm3あたり20万個にまで達しました(図2B)。深度約400 m・50℃以上の深部環境では、1 cm3あたりの定量限界値を上回る数百細胞程度の微生物群が散逸的に検出されました。75-90℃にかけての厚さ200 mの区間では、再び内生胞子の密度が増加し、85℃において1 cm3あたり120万個にまで達しました(図2B)。また、深度570-633 m・70℃付近とプレート境界断層下部の829-1021 m・90-110℃の区間では、通常の微生物細胞も内生胞子も検出されない非生命環境に近いゾーンが認められました。しかし、1021 mを超えた堆積物―基盤岩境界までの深度区間(1021-1180 m・110-120℃)では、定量限界値(1 cm3あたり16細胞)を上回る数の微生物細胞が検出され、深度が増加するにつれて細胞数が増加する傾向が認められました(図2A)。
一方、深度約730 m・80-85℃付近には、基盤岩から堆積物にむけて供給される硫酸と堆積物中のメタンの濃度が交差する硫酸―メタン境界域が認められました(図3A)。この深度より上部の堆積物では、微生物起源ガスを示す軽い炭素同位体比(平均 −61.3±3.0‰、※6)を持つメタンが存在し(図3B)、比較的高いメタン生成活性が認められました(図3D)。また、それより深い堆積物では、炭素同位体比の値が重い低濃度の熱分解起源のメタンが存在し、メタン生成活性も極めて低いか検出限界以下でした。また、80-85℃の高温の堆積物環境において、メタンの炭素同位体が局所的に重くなる傾向が認められました(図3B)。これは、メタンを硫酸還元などにより酸化する嫌気的メタン酸化反応を担う微生物生態系が存在することを示唆しています。さらに、細胞数が増加する深度1021-1180 m・110-120℃の区間では、再びメタンの炭素同位体組成が軽くなる傾向が認められると共に、有意なメタン生成活性が認められました(図3B,D)。
大変興味深いことに、深度600 m・70℃以上の深部堆積物では、メタン濃度や内生胞子密度の減少と対照的に、堆積物の間隙水中に含まれる酢酸の濃度が10 mM(600 mより浅い深度の堆積物に含まれる酢酸の量の400倍以上に相当)を上回る高濃度まで上昇している現象が認められました(図3C)。深度600-1000 mの区間では、酢酸の濃度と炭素同位体比がほぼ一定の値を示しました(平均9.2±2.4 mM、−18.8±0.5‰)。これは、有機物の分解により生成された酢酸の量に対して、微生物が消費する酢酸の量が極めて少ない(または、微生物により消費されていない)ことを示しています。しかし、1030 mを超える深度では、酢酸の濃度が深くなるにつれて減少し、酢酸の炭素同位体比の値も−7.9‰まで重くなる傾向が認められました(図3C)。この現象は、100℃を超える高温の堆積物―基盤岩境界域において、微生物がメタン生成や硫酸還元と共役した酢酸酸化反応を行い、軽い炭素同位体比を持つ酢酸を消費してエネルギーを得たためと考えられます(図4)。
5. 考察と今後の展望
上記の微生物学的、地球化学的、地質学的研究データは、表層堆積物に生息している多くの海底下微生物は温度に感受性がある好冷性〜常温性の微生物群集であり、40-50℃に存続温度限界があることが示されました。また、45-85℃の堆積物には、その温度に生育温度上限を持つ低密度の好熱性微生物が局所的に存在し、嫌気的メタン酸化反応などの物質循環に寄与していることが明らかとなりました。さらに、それらの微生物群集の一部は、存続限界温度(40-50℃と70℃付近)に近づくと内生胞子を形成するなどの生存戦略をとることが明らかになりました。一方、70℃前後や90-110℃の堆積物には生命シグナルに乏しい区間が存在していました。これは、プレート境界断層と基盤岩に挟まれた堆積物が、局所的に高い間隙圧と弱い力学強度を持ち(※7)、過去に温度の高い流体が間欠的に供給されたことで(※8)、微生物細胞が死滅または存続できない環境が生じている可能性が考えられます。しかし、110-120℃までの堆積物―基盤岩の境界域では、常温性微生物や好熱性微生物の生息域とは非連続的に、基盤岩から供給される深部流体(熱水)に含まれる化学成分や、堆積物に含まれる酢酸を栄養・エネルギー基質として消費する超好熱性微生物生態系の存在が示唆されました(図2C,5)。
これらの研究の成果は、宇宙において唯一生命が確認されている地球の内部環境において、温度をはじめとする物理的、化学的な因子の変化に対して、生命と生態系がどのように適応・存続し、その実態がどのように生命と地球惑星の共進化プロセスや物質循環に寄与しているのかを理解する上で、極めて重要な知見です。また、本研究では深度1021-1180 m・110-120℃の地質環境に超好熱性微生物の存在を示す生命シグナルが確認されました。本成果は、海底下深部の生命生息可能条件として、温度や地質学的な構造、栄養・エネルギーの確保が極めて重要な要因であることを示すのみならず、この惑星における海底下生命圏の空間規模やその限界が、120℃を上回る海洋地殻や上部マントルにまで達する可能性を示しています。
【補足説明】
- ※1
- コア試料:
掘削などによって採取される柱状の地質試料。
- ※2
- 国際深海科学掘削計画(IODP: International Ocean Discovery Program):
平成25年(2013年)10月から開始された多国間科学研究協力プロジェクト。日本(地球深部探査船「ちきゅう」)、アメリカ(ジョイデス・レゾリューション号)、ヨーロッパ(特定任務掘削船)がそれぞれ提供する掘削船を用いて深海底を掘削することにより、地球環境変動、地球内部構造、地殻内生命圏等の解明を目的とした研究を推進する。
- ※3
- 高知コアセンター:
高知大学と海洋研究開発機構が共同運営する掘削コア試料の中核的な分析研究拠点。科学掘削により採取された世界各地のコア試料を保管・管理する「世界三大コア保管施設」の一つ(高知コアセンターの他には、米国・テキサスA&M大学とドイツ・ブレーメン大学に類似のコア保管施設がある)。
- ※4
- 内生胞子:
バクテリア(真正細菌)の分類群の一つであるファーミキューテス門(Firmicutes)に属する微生物がもつ休眠状の細胞形態の一つ。生育限界に近い温度や、栄養が欠乏し飢餓状態になるなどのストレス条件下において、細胞内に耐久性の高い構造を形成する。本研究では、内生胞子に特異的に含まれる成分であるジピコリン酸の量を測定することで、堆積物に含まれる内生胞子の数を推定した。
- ※5
- プレート境界断層(デコルマ帯):
南海トラフにおいて、フィリピン海プレートが日本列島の下に沈み込む際に圧縮を受け、陸側に付加される堆積物と沈み込む堆積物との間に水平方向に発達する力学的な境界面。付加体が形成される沈み込み帯外縁部から初期形成がはじまり、深部流体の流れや応力の蓄積・解放に直接的に関係することから、海溝型地震の発生メカニズムを理解する上で極めて重要な地質構造とされる。
- ※6
- 微生物起源ガスを示す軽い炭素同位体比:
自然界の炭素原子(C)には、質量が異なる3種類の同位体(12C、13C、14C)が存在している。その内、質量数12の炭素(12C)が99%を占め、残りの1%に質量数13の安定同位体炭素(13C)と質量数14の放射性同位体炭素(14C)が含まれる。一般的に、(微)生物が自然界の炭素化合物を代謝する際に、質量の軽い分子を選択的に用いる傾向がある。それを、同位体分別効果という。微生物起源ガスは、微生物が軽い炭素(12C)を含む二酸化炭素などの基質からメタンを生産するため、12Cの比率が高く13Cの比率が低い「軽い」炭素同位体比を示す傾向がある。
- ※7
- 高い流体圧と弱い力学強度:
IODP第370次研究航海により、沈み込み帯先端部におけるプレート境界断層と基盤岩との間の区間(深度800-1050 m)には、その深度における静水圧より高い間隙圧を持つ堆積物環境(高間隙圧流体場)が存在し、その力学的強度が弱いことが明らかとなっている(Hamada et al., Progress in Earth and Planetary Science, 5, 70, 2018を参照)。
- ※8
- 過去に温度の高い流体が間欠的に供給されたこと:
IODP第370次研究航海により、プレート境界断層下部より得られた堆積物コア試料の亀裂に、熱水性の二次鉱物であるバライト(硫酸バリウム:BaSO4)やアンハイドライト(硫酸カルシウム:CaSO4)の沈殿が認められている。それらの鉱物は、約145-220℃の熱水が間欠的に供給され、温度の低下と共に結晶として析出したものであることが明らかになっている(Tsang et al., Marine and Petroleum Geology, 112, 104080, 2020を参照)。
- ※9
- ギブス自由エネルギー:
ある化学反応を一定の条件で想定した場合に、その化学ポテンシャル(熱化学的な平衡状態に至る自発的な振る舞い)は基質1分子あたりのギブス自由エネルギーで表すことができる。微生物にとってエネルギー源となる基質(反応物)のギブス自由エネルギーが減少すると、その代謝反応は生体内で自発的に進むと考えられる。
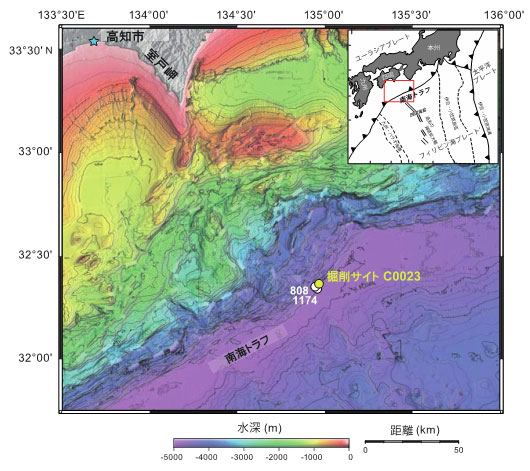
図1. IODP第370次研究航海で実施された高知県室戸岬沖の南海トラフ沈み込み帯先端部に位置する掘削地点(サイトC0023)。過去に、米国の掘削船ジョイデス・レゾリューション号を用いた国際深海掘削計画(ODP)第131次研究航海(サイト808)と第190・196次研究航海(サイト1174)が実施され、詳細な地質構造や周辺域に比べて地殻熱流量が高いことが報告されている。
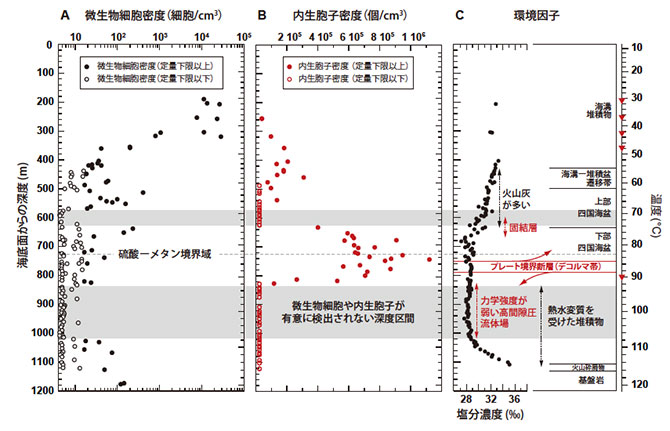
図2. IODP第370次研究航海で掘削地点C0023から採取された堆積物コア試料に含まれる微生物細胞と内生胞子の密度と環境因子の鉛直プロファイル。(A)蛍光色素(SYBR Green I)で染色した微生物細胞の密度プロファイル。定量下限値よりも高い細胞密度のプロットを●、低いプロットを○で示す。(B)内生胞子に特異的なバイオマーカーであるジピコリン酸の定量分析から推定される内生胞子の密度プロファイル。(C)堆積物の間隙水中の塩分濃度、温度、堆積学的特徴の概略図。基盤岩に向けての塩分濃度の上昇は、基盤岩(玄武岩)の変質の影響を受けた流体が堆積物側に供給されていることを示している。灰色の区間は、蛍光染色による微生物細胞とジピコリン酸による内生胞子の両方が有意に検出されなかった深度(570-633 mと829-1021 m)を示している。
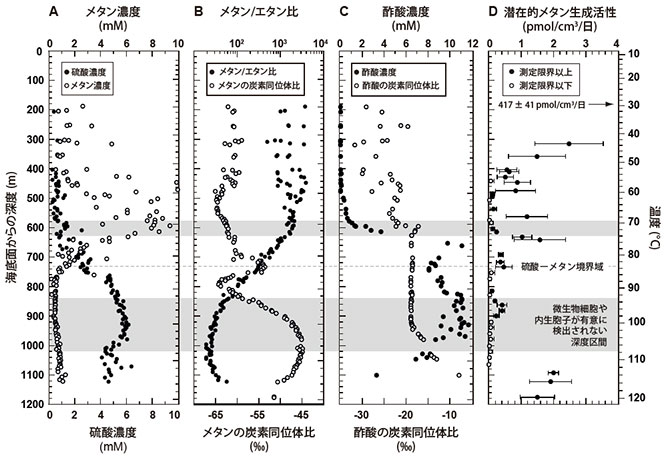
図3. 掘削地点C0023におけるガス・間隙水の地球化学的特性とメタン生成活性の深度プロファイル。(A)硫酸(●)とメタン(○)の濃度。(B)メタン/エタン比(●)とメタンの炭素同位体比(○)。(C)間隙水中の酢酸の濃度(●)と酢酸の炭素同位体比(○)。(D)14Cで標識されたCO2を放射性トレーサーとして用いた潜在的メタン生成活性(14CO2から14CH4への転換量に基づく水素資化性メタン生成反応の速度)。定量下限値(0.094 pmol/cm3/日)を超える測定値を●、それ以下で検出された測定値を○で示す。深度180 mでの潜在的メタン生成活性は417±41 pmol/cm3/日とスケール外の値を示した。これらの活性値は、深度360 mでは40℃、405-585 mの区間では60℃、604-775 mの区間では80℃、816 m以上の区間では95℃でインキュベーションを行い測定された。灰色の区間と温度は図2と同様である。
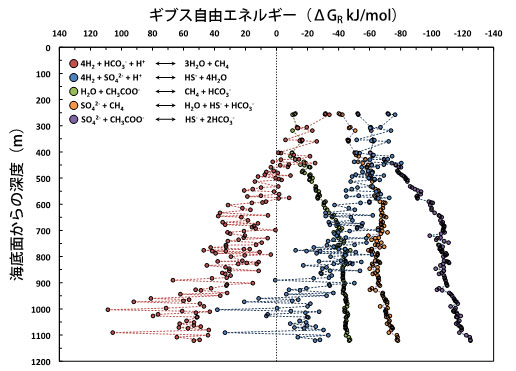
図4. 掘削地点C0023のガス及び間隙水化学成分の濃度に基づく微生物呼吸代謝反応のポテンシャルを示すギブス自由エネルギー(ΔGR、※9)の深度プロファイル。深度約400 m・50℃以上の堆積物では、CO2還元による水素資化性メタン生成反応(赤色)によりエネルギーを自発的に獲得することは困難である。また、水素資化性の硫酸還元反応(青色)も、深度が増加するにつれて基質(硫酸イオン)1モルあたりのギブス自由エネルギーが増大する傾向があり、反応は進みにくくなる。それらとは対照的に、酢酸開裂型のメタン生成反応(緑色)や、硫酸還元を伴う嫌気的メタン酸化反応(橙色)、酢酸を基質とする硫酸還元反応(紫色)については、深度が増加するにつれて反応基質のギブス自由エネルギーが減少し、それらの代謝様式によってエネルギーが獲得しやくすくなる傾向が認められた。
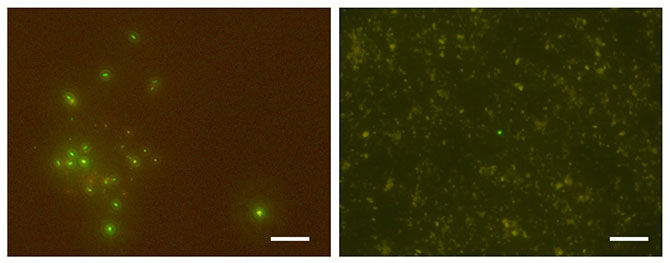
図5.掘削地点C0023より検出された微生物細胞の蛍光顕微鏡写真。細胞に含まれるDNAを緑色の蛍光試薬(SYBR Green I)で染色したもの。(左)深度652.0 m・76℃の堆積物コア試料(43R-3)から分取された微生物細胞。(右)深度1176.8 m・120℃の堆積物コア試料(112R-2)から検出された微生物細胞(写真中央に1細胞)。スケールは20マイクロメートル(1ミリメートルの1/50)を示す。
国際深海科学掘削計画(IODP)第370次研究航海「室戸沖限界生命圏掘削調査:T-リミット」(ダイジェスト版紹介ビデオ)
- (本研究について)
- 国立研究開発法人海洋研究開発機構
- 研究プラットフォーム運用開発部門 マントル掘削プロモーション室
室長 稲垣史生 - 超先鋭研究開発部門 高知コア研究所 地球微生物学研究グループ
主任研究員 諸野祐樹 - 国立大学法人東京大学 地震研究所 附属地震火山情報センター
- 教授 木下正高
- (報道担当)
- 国立研究開発法人海洋研究開発機構
- 海洋科学技術戦略部 広報課
- 国立大学法人高知大学
- 総務部総務課広報係
- 国立研究開発法人産業技術総合研究所
- 広報部 報道室
- 国立大学法人京都大学
- 総務部広報課 国際広報室
- 国立大学法人神戸大学
- 総務部 広報課